Caratterizzazione dei suoni di corteggiamento delle specie del sottogruppo fasciola (Diptera, Drosophilidae, gruppo Drosophila repleta): analisi interspecifiche e interpopolazionali
Traduzione automatica
L'articolo originale è scritto in lingua EN (link per leggerlo) .
Abstract
L'obiettivo di questo lavoro era caratterizzare il modello di canto di corteggiamento maschile di varie specie del sottogruppo fasciola e determinare il livello di variazione sia all'interno che tra le specie. I parametri analizzati erano l'intervallo intrapulsato (PI), l'intervallo interpulsato (IPI) e la frequenza intrapulsata (IF). Sono state analizzate sei diverse specie: D. coroica (tre popolazioni), D. ellisoni, D. fascioloides, D. moju, D. onca e D. rosinae (una popolazione ciascuna). Ci sono state differenze significative tra le sei specie per questi tre parametri del canto di corteggiamento. L'IPI era il parametro più variabile tra queste specie, suggerendo che questo parametro è importante per la discriminazione femminile. Quattro diverse ipotesi potrebbero spiegare questa variazione: 1. diverse pressioni selettive con assenza di flusso genico; 2. selezione sessuale intraspecifica; 3. effetti simpatici sull'evoluzione del canto; e 4. deriva genetica. Il PI era l'unico parametro che era significativamente diverso tra le tre popolazioni di D. coroica. Bassa variabilità tra le popolazioni all'interno della stessa specie era già stata osservata per altri sottogruppi e potrebbe essere spiegata dalle seguenti ipotesi: forte selezione che agisce sui parametri del canto, flusso genico o recente colonizzazione da una fonte comune. Studi aggiuntivi sul canto di corteggiamento di altre specie del sottogruppo fasciola, così come per altri sottogruppi del gruppo repleta, e studi, utilizzando marcatori molecolari, che si concentrano sulla base genetica delle differenze tra queste specie nel canto di corteggiamento ci permetterebbero di valutare l'associazione tra canto di corteggiamento e isolamento sessuale in queste specie, e ci aiuterebbero anche a comprendere l'evoluzione di queste differenze comportamentali.
Introduzione
Suoni di corteggiamento
Il comportamento di corteggiamento delle mosche del genere Drosophila è importante per il riconoscimento delle specie. Dopo che Shorey (1962) ha dimostrato che i maschi di D. melanogaster emettevano suoni durante il corteggiamento sessuale, sono stati condotti molti studi con i vari gruppi e sottogruppi del genere Drosophila, descrivendo la natura specifica dei tipi di suono. Questi studi hanno fornito prove che il suono ha una funzione sia come stimolante sessuale che nell'isolamento sessuale. Questi suoni variano significativamente tra le varie specie dei gruppi affinis (Chang & Miller, 1978), melanogaster (Cowling & Burnet, 1981), virilis (Hoikkala et al., 1982; Hoikkala & Isoherranen, 1997), mercatorum (Ikeda & Maruo, 1982), repleta (Ewing & Miyan, 1986), auraria (Tomaru & Oguma, 1994), planitibia (Hoikkala et al., 1994), willistoni (Ritchie & Gleason, 1995), quinaria (Neems et al., 1997) e obscura (Noor & Aquadro, 1998).
Le differenze interspecifiche nei suoni di corteggiamento di Drosophila sono caratterizzate da uno o più parametri sonori come: intervallo interpulsato (IPI) che è il tempo dall'inizio di un impulso all'inizio del successivo; intervallo intrapulsato (PI) che è la durata dell'impulso; frequenza intrapulsata (IF) che è acusticamente la frequenza più forte all'interno dell'impulso; numero di impulsi per serie; e durata di ciascun intervallo all'interno di una serie (Hoikkala & Lumme, 1987; Wheeler et al., 1988). Anche se non è stato stabilito con certezza quali parametri sonori conferiscano specificità, ci sono evidenze che l'IPI sia importante. Secondo Ritchie & Kyriacou (1996), l'IPI dei suoni di corteggiamento di Drosophila varia molto poco all'interno delle popolazioni naturali ed è importante per il successo riproduttivo maschile.
Tuttavia, è chiaro che molte specie di Drosophila producono suoni complessi con varie caratteristiche, una o più delle quali possono essere utili per identificare le specie. Questa specificità nei suoni di corteggiamento di Drosophila può essere spiegata dalla selezione per la discriminazione tra le specie (Tomaru & Oguma, 1994) dei diversi tipi di suoni, che potrebbero essere evoluti attraverso la selezione sessuale intraspecifica (Ritchie & Gleason, 1995). Inoltre, i diversi tipi di suoni presenti nelle varie specie di Drosophila potrebbero semplicemente riflettere le divergenze filogenetiche.
Il sottogruppo fasciola
Le specie del sottogruppo fasciola si trovano nelle foreste umide, principalmente in quelle dell'America Centrale e Meridionale e nelle isole dei Caraibi. In Brasile sono state raccolte dalla foresta pluviale atlantica, così come da foreste negli stati di São Paulo, Paraná, Mato Grosso do Sul e Bahia (Vilela, 1983; Vilela et al., 1983; Tidon-Sklorz & Sene, 1992).
La maggior parte del lavoro su questo sottogruppo è stata di natura citologica ed è stata svolta all'inizio degli anni '60, dimostrando che le specie di questo gruppo differiscono da tutte le altre del gruppo repleta per avere le inversioni omozigoti: 2o2, 2e3 e 2l3. Wasserman (1982) ha proposto un'ascendenza comune per i sottogruppi fasciola e mulleri, poiché condividono l'inversione 3c. Recentemente, tuttavia, Diniz (1998) ha dimostrato che l'inversione 3c del sottogruppo fasciola è diversa da quella trovata nel sottogruppo mulleri e ha proposto che il sottogruppo fasciola derivi direttamente dal primitivo I, e non possa più essere considerato legato al sottogruppo mulleri. Kuhn et al. (1995) hanno effettuato uno studio cariotipico di alcune specie del sottogruppo fasciola e hanno trovato variazioni interspecifiche nella dimensione del microcromosoma e nel numero di cromosomi.
Secondo Wasserman (1992), il fasciola sottogruppo consiste di nove specie: D. pictilis e D. pictura, formando il complesso pictilis; D. paraguttata; D. mojuoides e D. moju, formando il complesso moju; e D. fulvalineata, D. fasciola, D. coroica e D. fascioloides che non si trovano in alcun complesso. Vilela (1983), studiando la morfologia dei genitali maschili, ha determinato che la specie D. fascioloides studiata da Wasserman (1962) in realtà non apparteneva a questa specie, e l'ha rinominata D. ellisoni. Ha anche aggiunto le specie D. linearepleta e D. onca al sottogruppo fasciola e descritto D. carolinae, D. hermionae, D. ivai, D. querubimae, D. rosinae e D. senei come nuove specie, aumentando a 18 il numero di specie in questo sottogruppo.
In questo lavoro descriviamo i modelli sonori di corteggiamento di sei specie del sottogruppo fasciola: D. ellisoni, D. fascioloides, D. moju, D. onca, D. rosinae e D. coroica. Oltre alle analisi interspecifiche, abbiamo anche esaminato le differenze nei modelli sonori di tre diverse popolazioni di D. coroica.
Materiali e metodi
Stabilimento e mantenimento delle isoline
Le isoline sono state prodotte da femmine individuali già accoppiate in natura. Queste isoline sono state identificate attraverso un'analisi morfologica dei genitali dei maschi F1 (Vilela, 1983). Le larve di terzo stadio del sottogruppo fasciola normalmente tentano di strisciare fuori dal flacone di coltura. Per evitare questo problema, i flaconi di coltura sono stati aperti e posizionati su sabbia umida all'interno di bottiglie di plastica in modo che le larve potessero pupa nella sabbia. Le mosche eclodate sono state rimosse con un aspiratore e trasferite in flaconi riempiti con mezzo di coltura, dove sono maturate, si sono accoppiate e hanno deposto uova, riavviando il ciclo.
Registrazioni sonore
Mosche vergini recentemente eclodate sono state isolate in flaconi di coltura da 35 ml e lasciate per 7-10 giorni per consentire il tempo per la maturazione sessuale. Successivamente, un maschio e una femmina della stessa linea sono stati introdotti in una camera di registrazione sonora descritta da Sene & Manfrin (1998). Per registrare il suono, la camera contenente la coppia di mosche è stata posizionata sul diaframma di un microfono ultrasensibile, come descritto da Manfrin et al. (1997). Le mosche sono state osservate per un massimo di 10 minuti, o fino all'accoppiamento. Tutte le registrazioni sono state effettuate a 25 ± 1ºC.
Analisi del suono
I segnali elettromagnetici delle registrazioni analogiche sono stati convertiti in segnali digitali a una frequenza di 5 KHz, con un circuito convertitore analogico-digitale CAD 12/36-60 k (Lynx) e il programma AqDADOS 4 (Lynx).
I modelli sonori di corteggiamento di sei diverse specie del sottogruppo fasciola sono descritti nella Tabella 1. I modelli sonori dei maschi di D. coroica provenienti da tre diverse località sono stati anche analizzati per studiare le differenze interpopolazionali.
I parametri utilizzati per gli studi comparativi dei modelli sonori erano l'intervallo intrapulse (PI), l'intervallo interpulse (IPI) e la frequenza intrapulse (IF). I valori di PI e IPI sono stati ottenuti direttamente dalle figure. Le IF sono state ottenute con l'aiuto del software MATLAB (MathWorks, Inc.) utilizzando le Trasformate di Fourier Veloci (FFT). Questa analisi scompone il segnale sonoro in una serie di onde (seni e coseni) di diverse frequenze e ampiezze. L'IF di un suono è definita come la frequenza dell'onda di massima ampiezza nel segnale sonoro.
La FFT può analizzare suoni contenenti migliaia di punti per millisecondo, ma l'efficienza massima dell'analisi si ottiene limitando il numero di punti a potenze di base due. Per questo motivo, tutti gli intervalli utilizzati per l'analisi dell'IF sono stati selezionati in modo che avessero un numero di punti pari a potenze di base due. Pertanto, sono stati utilizzati intervalli di 64 punti per D. moju; 128 punti per D. rosinae e D. coroica; 256 punti per D. ellissoni; e 512 punti per D. onca e D. fascioloides. Sono state effettuate dieci misurazioni dei parametri PI, IPI e IF per ciascuno dei 10 maschi di ciascuna delle isoline.
Analisi statistica
Le linee descritte nella Tabella 1 sono state analizzate con l'obiettivo di indagare le differenze interspecifiche. L'isolato D96N74 è stato scelto come rappresentativo di D. coroica. Le tre linee (D96N74, D88M1 e J7C1) di D. coroica sono state testate separatamente per le differenze interpopolazionali. I valori dei parametri intervallo intrapulsato (PI), intervallo interpulsato (IPI) e frequenza intrapulsata (IF) sono stati analizzati separatamente mediante analisi della varianza (ONE-WAY ANOVA) a un livello di significatività di 0,05.
Risultati
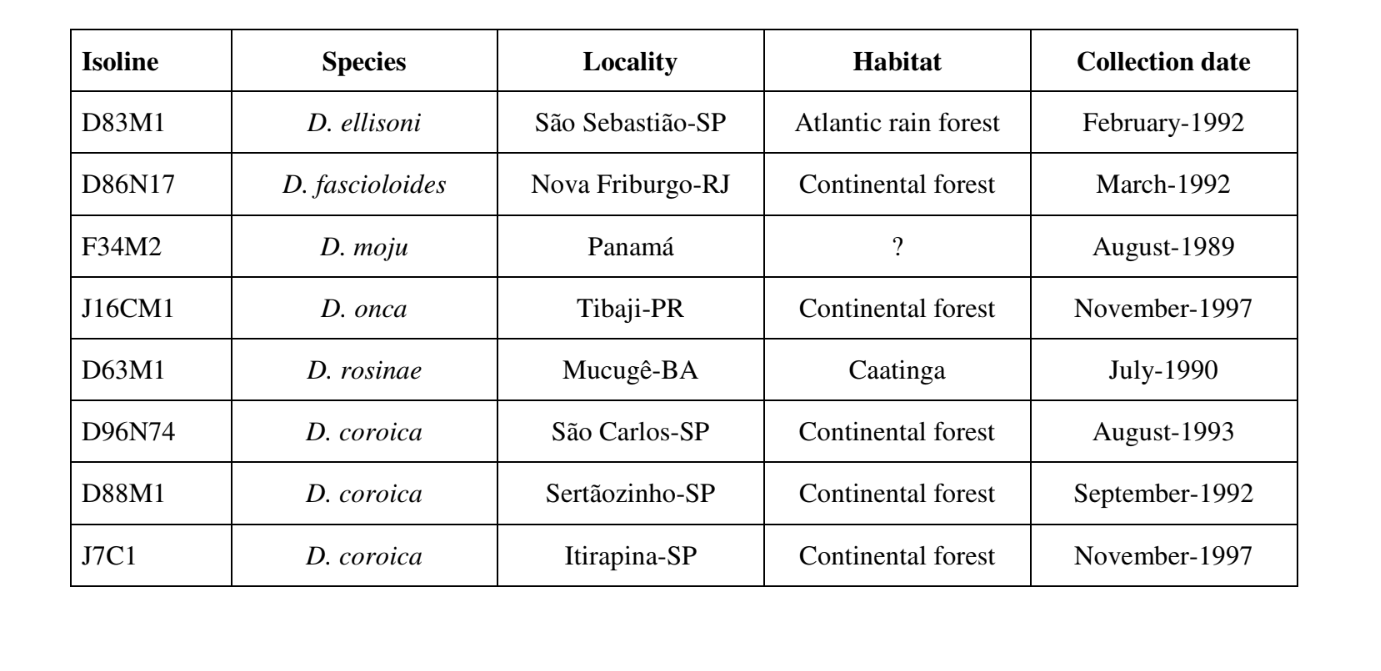
Le Figs. 1 a 6 mostrano i sonogrammi di base delle varie specie analizzate. L'isolato D96N74 è stato scelto per rappresentare la specie D. coroica. Questi sonogrammi e l'analisi dei impulsi sonori consentono una divisione dei suoni di corteggiamento in tre gruppi distinti: 1. suoni di corteggiamento delle specie D. onca, D. fascioloides e D. ellisoni, in cui ciascuna sequenza sonora è composta da una sequenza di brevi impulsi, seguita da un trill, simile a quello trovato in alcune specie del sottogruppo planitibia, denominato da Hoikkala et al. (1994) come frase canora. Nelle specie D. fascioloides e D. onca, la sequenza di impulsi è composta da quattro impulsi, seguiti da un trill relativamente lungo, mentre in D. ellisoni, ci sono solo due impulsi, seguiti da un trill più corto; 2. il tipo tipico del sonogramma di D. rosinae, con un suono di corteggiamento formato da impulsi semplici ma duplicati, un modello osservato anche da Ewing & Miyan (1986) in alcune specie del gruppo repleta; 3. un tipo basato sui suoni di corteggiamento delle specie D. moju e D. coroica, con impulsi brevi in sequenza, simile alla canzone a impulsi di D. melanogaster (Schilcher, 1976) e suono A di D. mercatorum (Ikeda & Maruo, 1982; Manfrin et al, 1997).
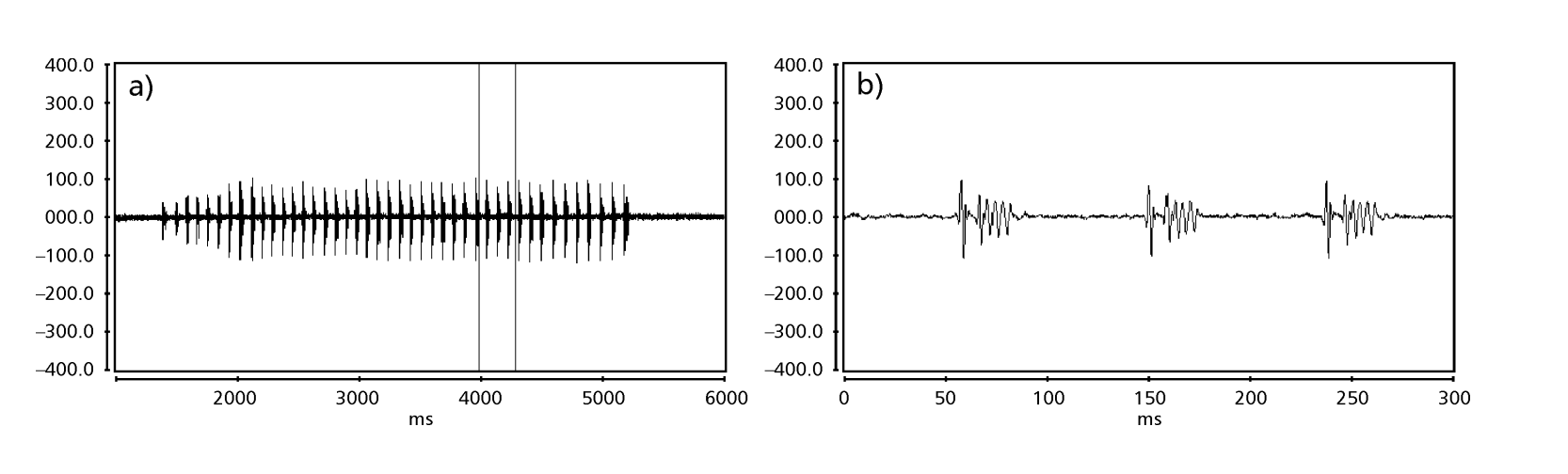
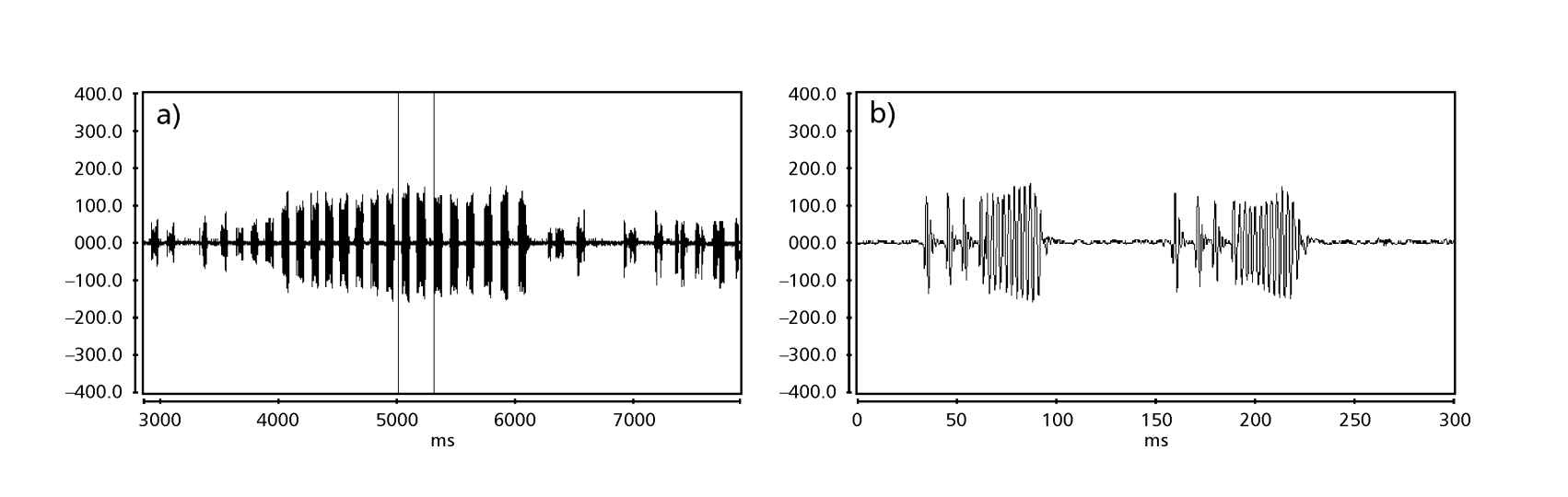
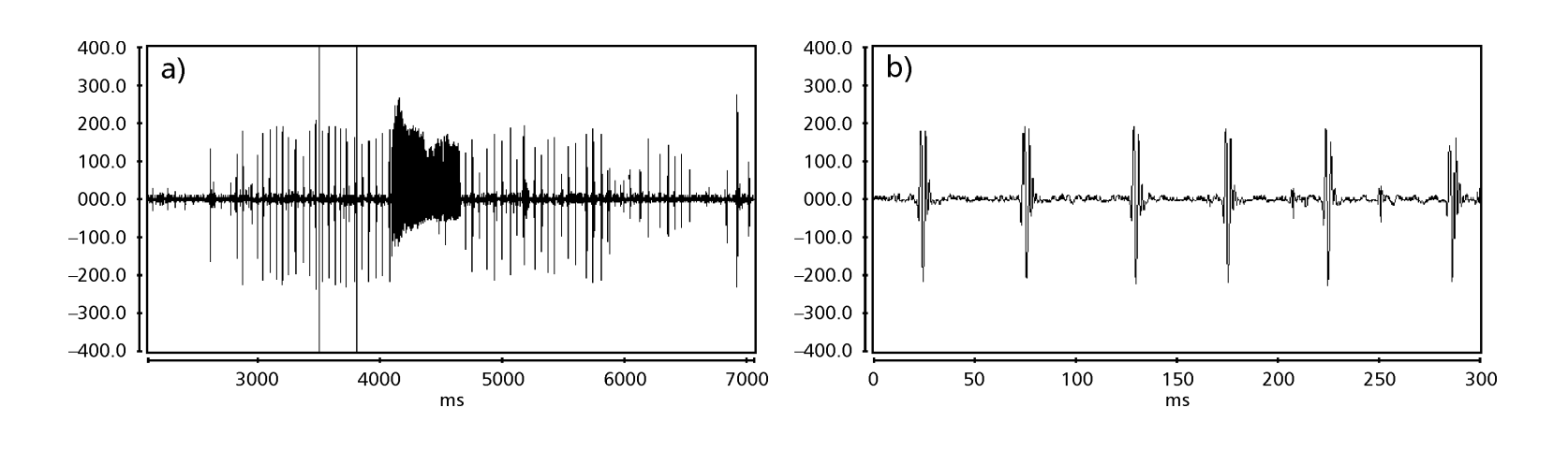
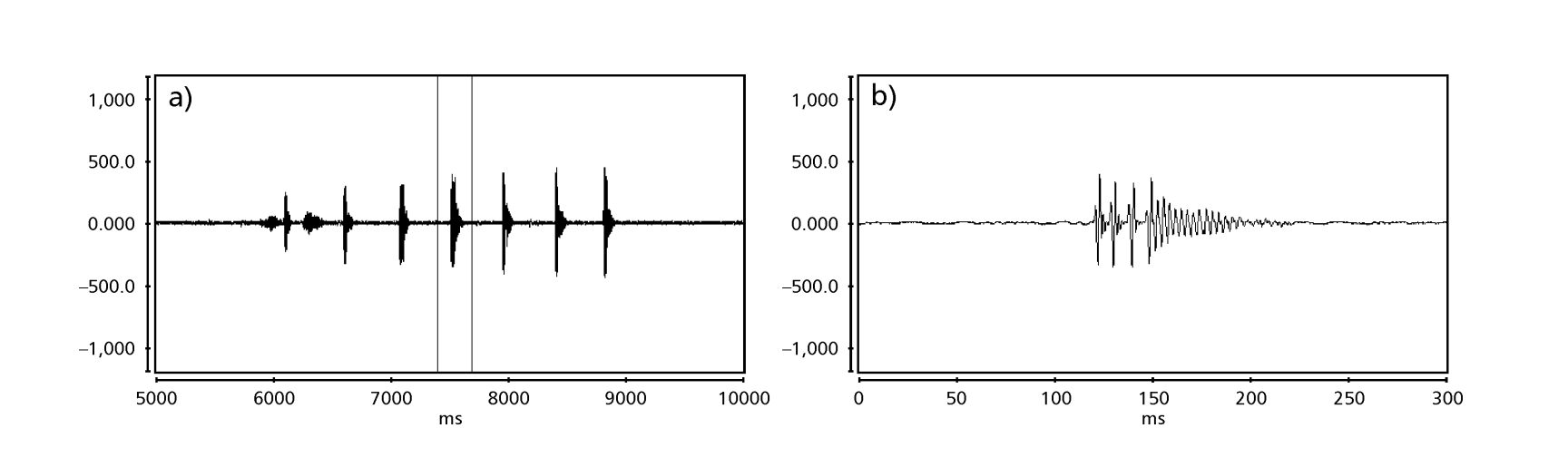
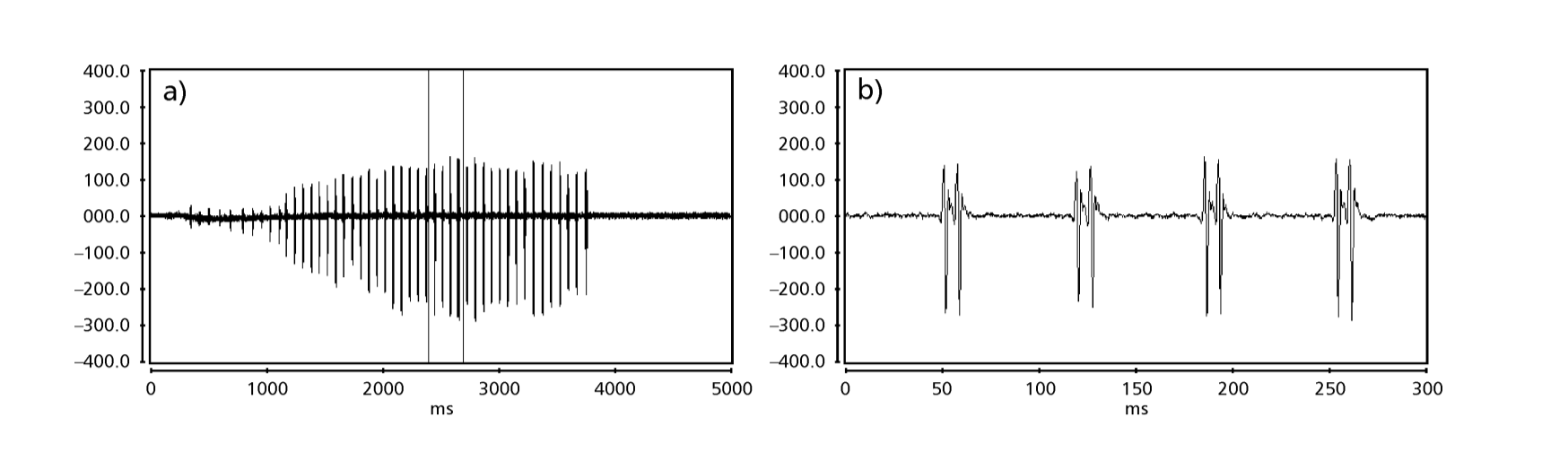
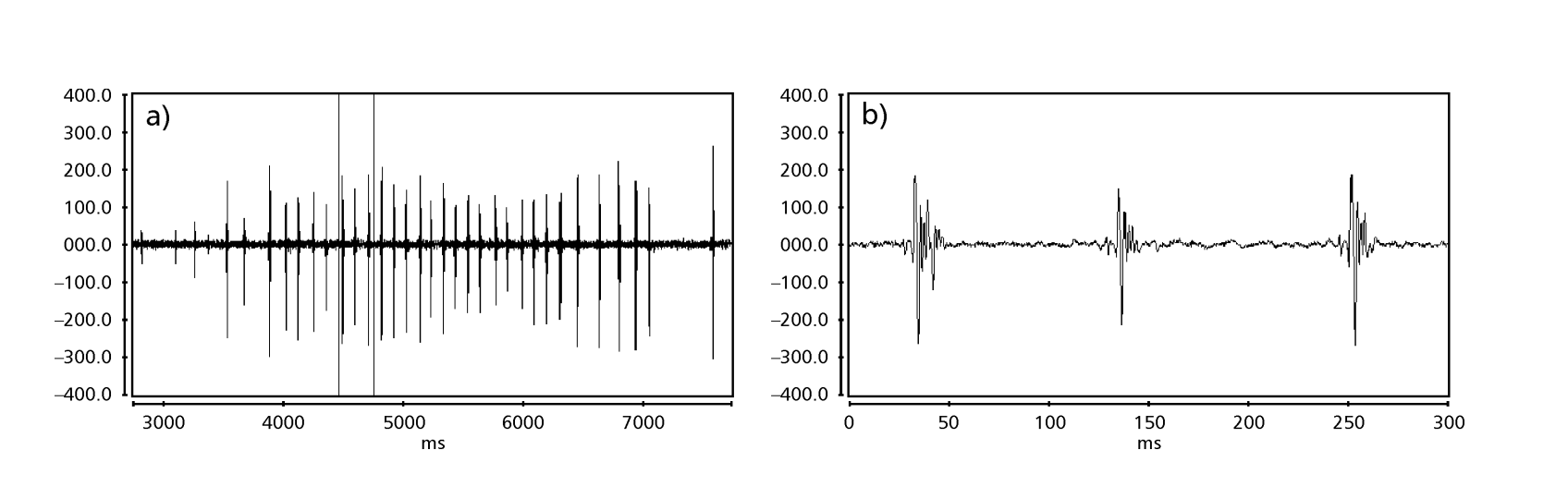
I sonogrammi di ciascuna delle specie analizzate erano una sequenza di un tipo di impulso sonoro. Questa scoperta è diversa da quelle degli studi effettuati sulle specie D. melanogaster (Schilcher, 1976), D. mercatorum (Ikeda & Maruo, 1982; Manfrin et al., 1997) e alcune specie del sottogruppo repleta (Ewing & Miyan, 1986), in cui sono stati trovati due diversi tipi di suoni.
La Tabella 2 mostra i valori medi per PI (intervallo intrapulsato), IPI (intervallo interpulsato) e IF (frequenza intrapulsata) delle isolinee descritte nella Tabella 1. Un confronto delle deviazioni standard (± sd) tra le varie specie mostra che D. ellisoni e D. fascioloides hanno la minore variazione per tutti e tre i parametri: PI (27.470 ± 2.73 e 69.859 ± 6.12), IPI (97.473 ± 5.76 e 125.757 ± 7.87) e IF (251.358 ± 19.36 e 362.901 ± 17.04), indicando che i suoni di corteggiamento di queste specie sono uniformi con poca variazione intraspecifica. D'altra parte, D. onca ha mostrato una variazione considerevole in PI (93.662 ± 31.44) e IPI (508.091 ± 73.06) e una variazione moderatamente alta in IF (289.260 ± 32.71) rispetto alle altre specie, indicando che le differenze in PI e IPI, sebbene grandi, non erano sufficienti a modificare significativamente la forma d'onda dell'impulso. Le deviazioni standard erano relativamente basse per PI (7.363 ± 7.36 e 14.673 ± 1.98) e IPI (50.859 ± 5.47 e 72.340 ± 5.68) in D. moju e D. rosinae, ma i valori di IF erano relativamente alti (412.501 ± 55.63 e 306.624 ± 57.80). In questo caso, sebbene la durata dell'impulso e il tempo da un impulso all'altro fossero relativamente uniformi all'interno di ciascuna di queste specie, la forma d'onda dell'impulso sonoro variava considerevolmente.
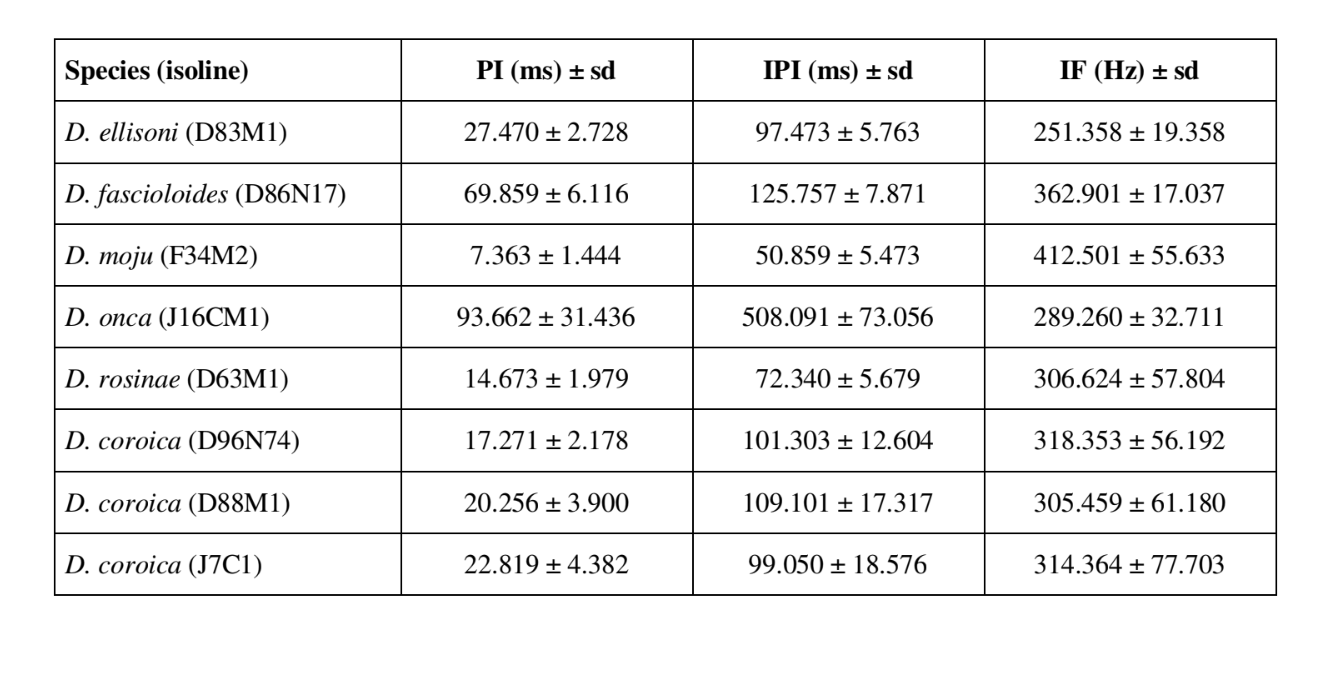
C'era relativamente poca variazione intrapopolazionale nei PI delle linee D. coroica D96N74, D88M1 e J7C1 (17.271 ± 2.18, 20.256 ± 3.90 e 22.819 ± 4.38, rispettivamente), variabilità intermedia nell'IPI (101.303 ± 12.60, 109.101 ± 17.32 e 99.050 ± 18.58) e alta variabilità nell'IF (318.353 ± 56.19, 305.459 ± 61.18 e 314.364 ± 77.70), indicando che anche con una bassa variazione intrapopolazionale nella lunghezza dell'impulso sonoro, c'era una considerevole variazione nella forma d'onda dell'impulso sonoro.
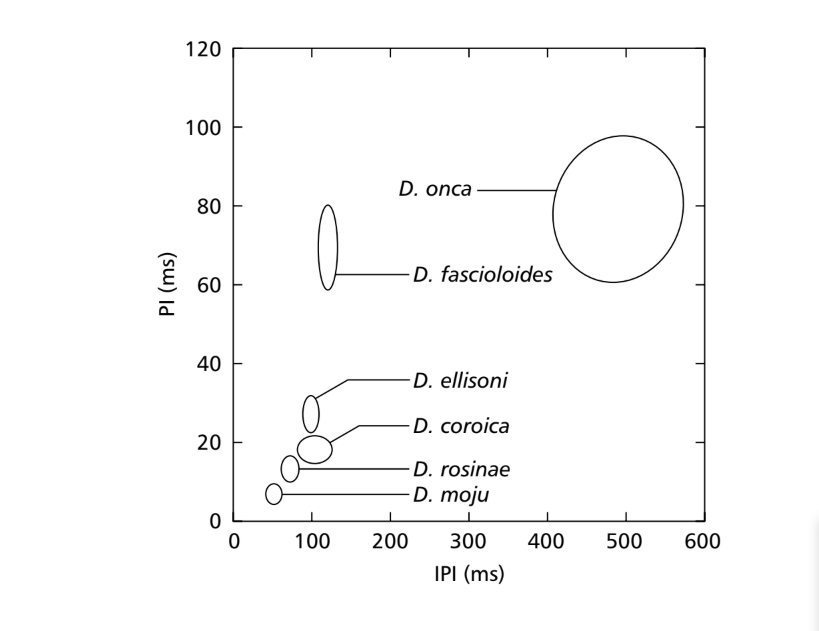
Quando i valori medi per i parametri PI e IPI sono stati confrontati, sono state trovate differenze tra le specie del sottogruppo fasciola per almeno uno di questi parametri. Queste differenze nei suoni di corteggiamento possono essere viste nella Fig. 7, in cui i valori per questi due parametri sono rappresentati sotto forma di ellissi, che includono il 75% dei valori per i parametri PI e IPI. Sono state trovate differenze significative nei valori di PI (ANOVA, F5,48 = 124.98, p < 0.001) e IPI (ANOVA, F5,48 = 949.35, p < 0.001) tra le specie. Le differenze interspecifiche in IF sono mostrate nella Fig. 8. Sono state trovate anche differenze significative nei valori di IF (ANOVA, F5,48 = 23.40, p < 0.001).
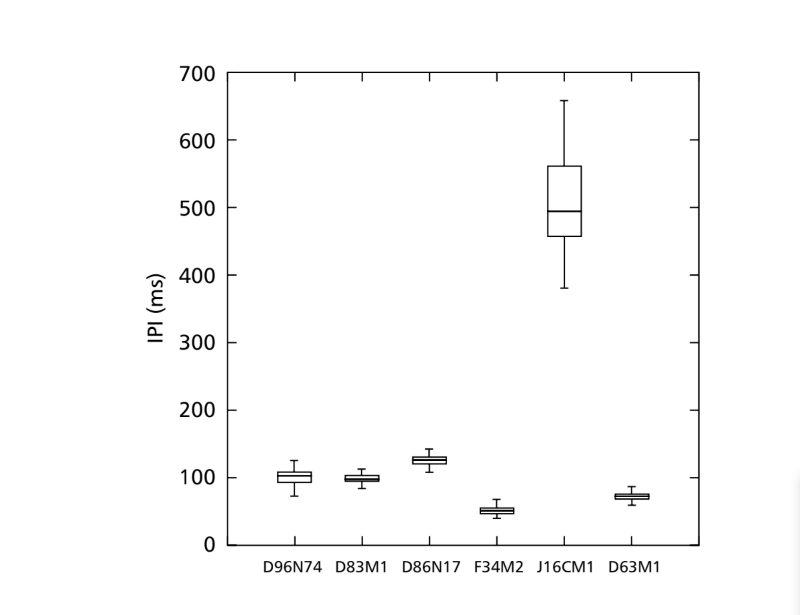
Le tre popolazioni di D. coroica differivano significativamente nel PI (ANOVA, F2,24 = 8.75, p < 0.05) ma non nell'IPI (ANOVA, F2,24 = 1.72, p > 0.10) e IF (ANOVA, F2,24 = 0.15, p > 0.25).
Discussione
Hoikkala et al. (1994) nella loro analisi del suono di corteggiamento di varie specie del sottogruppo planitibia, hanno scoperto che è comune trovare gli stessi tipi di suoni in specie filogeneticamente vicine. Inoltre, Hoikkala & Kaneshiro (1993) hanno osservato che alcune specie strettamente correlate in questo sottogruppo, che sono simpatiche, utilizzano diversi tipi di suoni nel corteggiamento.
Noor & Aquadro (1998) hanno trovato differenze significative nei suoni di corteggiamento in due specie strettamente correlate del sottogruppo obscura: D. pseudoobscura e D. persimilis. Alcuni esperimenti che coinvolgono incroci tra queste due specie hanno indicato che le femmine di D. pseudoobscura probabilmente hanno aumentato la loro discriminazione contro i maschi di D. persimilis nelle popolazioni simpatiche. Questo potrebbe essere avvenuto attraverso la selezione contro la produzione di ibridi mal adattati, risultando in un'alterazione del suono di corteggiamento in queste popolazioni simpatiche.
Ewing & Miyan (1986), lavorando con specie del gruppo repleta , non hanno trovato alcuna relazione tra una filogenesi basata su dati citologici e un'altra basata su dati sonori. Tuttavia, alcune delle caratteristiche che la maggior parte delle specie studiate ha in comune e che si trovano anche in specie di altri gruppi ci permettono di proporre un suono di corteggiamento ancestrale per il gruppo repleta . Questo suono sarebbe stato composto da due componenti distinte: il suono iniziale A, costituito da un massimo di 10 impulsi monociclici brevi e regolari, con un intervallo interimpulso di 8-15 ms; e il suono B, composto da una serie di impulsi regolari e monociclici, ma con un numero maggiore di impulsi e un intervallo interimpulso più lungo alla fine del corteggiamento. Durante l'evoluzione del gruppo repleta , alcune specie avrebbero perso il suono A, e altre, il suono B, e, in molte, il suono B sarebbe diventato meno regolare e più complesso.
Basandoci su queste supposizioni e osservando i sonogrammi ottenuti per le sei specie del sottogruppo fasciola in questo studio, possiamo inferire che in tutte queste specie c'è stata una perdita del suono A, poiché gli intervalli interimpulso sono sempre molto più lunghi di 15 ms. D. coroica ha impulsi irregolari, con un intervallo interimpulso medio di 101.3 ms, mentre i suoni di corteggiamento di D. moju sono composti da impulsi più regolari, con un intervallo interimpulso medio di
50.9 ms. D. rosinae ha un sonogramma composto da impulsi duplicati, un tipo di suono trovato anche da Ewing & Miyan (1986), con un intervallo medio tra impulsi di 72.3 ms. D. ellisoni, D. fascioloides e D. onca hanno suoni di corteggiamento composti da impulsi policiclici, probabilmente a causa di una fusione dei suoni A e B, con intervalli medi tra impulsi di 97.4, 125.7 e 508.0 ms, rispettivamente. Queste somiglianze nei tipi di suoni in queste tre specie non sono strettamente correlate ai dati citologici, tranne per D. ellisoni e D. fascioloides che sono stati collocati vicini l'uno all'altro in una filogenesi basata su caratteri citologici, hanno due fusioni cromosomiche in comune: 2-4F e 3-5F (Diniz, 1998). Come nel caso dei caratteri morfologici, ci si aspetta una tendenza affinché specie strettamente correlate condividano alcuni parametri sonori. Tuttavia, anche quando le relazioni filogenetiche tra le specie sono note, le tendenze evolutive basate su caratteri sonori non sono state ben stabilite.
Anche se il suono di corteggiamento non è un esempio di comportamento circadiano, la ricerca ha collegato questo comportamento all'espressione del gene period (per) situato sul cromosoma X e ben caratterizzato a livello molecolare. Le mutazioni in questo gene possono alterare il ritmo circadiano di Drosophila. Un'analisi genetica condotta da Konopka et al. (1996) ha mostrato che il ritmo sonoro dell'intervallo interpulsato è anch'esso influenzato da mutazioni nel gene per. Kyriacou & Hall (1980) hanno riportato una modulazione ritmica dell'intervallo interpulsato con periodi specifici in D. melanogaster e D. simulans di 55 e 35 secondi rispettivamente. Queste differenze sono state dimostrate essere correlate ad alcune variazioni nel gene per. Le mutazioni in questo gene, che sono state inizialmente studiate da Konopka & Benzer (1971) in un'analisi dei ritmi circadiani anomali, hanno provocato effetti corrispondenti nei ritmi degli intervalli interpulsati dei suoni di corteggiamento. La mutazione perShort (perS), con un ritmo circadiano di circa 19-20 ore, aveva un periodo medio di 41,5 secondi; la mutazione perLong (perL) il cui ritmo circadiano di 28-30 ore aveva un periodo medio di 82,1 secondi; e la mutazione per0 non aveva alcun ritmo di intervallo interpulsato.
In questa ricerca abbiamo trovato differenze significative nei parametri intervallo intrapulsato, intervallo interpulsato e frequenza intrapulsata nelle specie di fasciola analizzate. Tra questi parametri, l'intervallo interpulsato è stato quello che ha mostrato maggiori differenze, con poca sovrapposizione e poca variazione intraspecifica, suggerendo che questo parametro potrebbe far parte di un sistema di riconoscimento delle specie. Questo alto livello di differenziazione nei suoni maschili è coerente con il consenso generale che il comportamento di corteggiamento possa differire più dei caratteri morfologici o di altre caratteristiche nelle specie correlate. Tuttavia, al momento è impossibile sapere la direzione dell'evoluzione di questi diversi tipi di suoni nel sottogruppo fasciola. Studi genetici hanno dimostrato che le differenze interspecifiche negli intervalli interpulsati sono frequentemente dovute a fattori additivi e poligenici. Queste differenze possono anche essere spiegate dall'adattamento a diversi ambienti in assenza di flusso genico. È noto che ci sono stati cicli alternati tra freddo-secco e caldo-umido nel periodo quaternario. In un ciclo climatico freddo-secco, la vegetazione adattata alle condizioni secche si è espansa, mentre la foresta si è ritirata. Durante il successivo ciclo caldo-umido la situazione si è invertita (Ab’ Saber, 1977; Vanzolini, 1981). Si crede che le mosche abbiano seguito questi processi di riduzione ed espansione della foresta, il che avrebbe comportato lunghi periodi di isolamento geografico durante i cicli freddo-secco e alcune aree di simpatria durante i cicli caldo-umido.
Sebbene l'intervallo interpulsato possa variare ampiamente tra le diverse specie del genere Drosophila, generalmente ciò non viene osservato quando si analizzano popolazioni della stessa specie (Costa et al., 2000; Ritchie et al., 1994). Le tre popolazioni analizzate di D. coroica presentavano differenze significative solo nell'intervallo intrapulsato, senza differenze nell'intervallo interpulsato e nella frequenza intrapulsata. È possibile che una forte pressione selettiva stia agendo sull'intervallo interpulsato e sulla frequenza intrapulsata. Questa bassa variabilità potrebbe anche verificarsi se popolazioni geograficamente distinte fossero geneticamente omogenee a causa di un flusso genico costante, o se un antenato comune avesse recentemente colonizzato queste popolazioni. Tuttavia, i dati PI mostrano che non è così.
Si sa poco sui meccanismi genetici coinvolti nei processi di speciazione, tuttavia è prima necessario comprendere la genetica e l'evoluzione dei meccanismi che ostacolano il flusso genico. I ibridi sterili e instabili sono considerati forti barriere al flusso genico, ma questi tipi di meccanismi postzigotici possono comportare alti costi sotto forma di gameti ed energia. Pertanto, le differenze nei suoni di corteggiamento possono essere un importante meccanismo isolante, poiché limiterebbero o prevenirebbero tali perdite (Welbergen et al., 1992). L'analisi di questi segnali, che sono importanti per il successo dell'accoppiamento, è di notevole interesse per la biologia evolutiva e la conoscenza dell'architettura dei suoni potrebbe aiutare a dimostrare vari modelli di selezione sessuale e speciazione.
Autori: Costa, C. T. A. e Sene, F. M.
Riferimenti:
- Ab’ Saber, A. N., 1977, Spazi occupati dall'espansione dei climi secchi dell'America del Sud, in occasione dei periodi glaciali quaternari. Paleoclimas, 3: 1-19.
- Chang, H. C. & Miller, D. D., 1978, Suoni di corteggiamento e accoppiamento in specie del Drosophila affinis sottogruppo. Evoluzione, 32: 540-550.
- Costa, C. T. A., Kuhn, G. C. S. & Sene, F. M., 2000, Bassa variazione del canto di corteggiamento nelle popolazioni brasiliane del Sud e Sud-est di D. meridionalis (Diptera, Drosophilidae). Rev. Brasil. Biol., 60: 53-61.
- Cowling, D. E. & Burnet, B., 1981, Canti di corteggiamento e controllo genetico delle caratteristiche acustiche in specie sorelle del Drosophila melanogaster sottogruppo. Anim. Behav., 29: 924-935.
- Diniz, N. M., 1998, Filogenia cromosomica di specie del sottogruppo fasciola del gruppo repleta genere Drosophila. Tesi di Dottorato, Facoltà di Medicina di Ribeirão Preto dell'Università di São Paulo.
- Ewing, A. W. & Miyan, J. A., 1986, Selezione sessuale, isolamento sessuale e l'evoluzione del canto nel gruppo di specie Drosophila repleta. Anim. Behav., 34: 421-429.
- Hoikkala, A. & Lumme, H., 1987, La base genetica dell'evoluzione dei suoni di corteggiamento maschili del gruppo Drosophila virilis. Evoluzione, 41: 827-845.
- Hoikkala, A. & Kaneshiro, K. Y., 1993, Cambiamento nella sequenza segnale-risposta responsabile per l'isolamento asimmetrico tra Drosophila planitibia e D. silvestris. Proc. Natn,. Acad. Sci., 90: 5813-5817.
- Hoikkala, A. & Isoherranen, E., 1997, Variazione e ripetibilità dei caratteri del canto di corteggiamento tra maschi di Drosophila montana e D. littoralis catturati in natura e allevati in laboratorio (Diptera-Drosophilidae). Jornal of Insect Behavior, 10: 193-202.
- Hoikkala, A., Lakovaara, S. & Romppainen, E., 1982, Comportamento di accoppiamento e suoni di corteggiamento maschili nel gruppo Drosophila virilis. In: Lakovaara, S. (ed.), Progressi nella genetica, sviluppo ed evoluzione di Drosophila. Plenum, New York.
- Hoikkala, A., Kenneth, Y. & Hoy, R. H., 1994, Canti di corteggiamento delle specie del sottogruppo Drosophila planitibia con ali a forma di immagine. Anim. Behav., 47: 1363-1374.
- Ikeda, H. & Maruo, O., 1982, Selezione direzionale per la frequenza di ripetizione del suono di corteggiamento e risposte correlate che si verificano in diversi caratteri in Drosophila mercatorum. Jnp. J. Genet., 57: 241-258.
- Konopka, R. J. & Benzer, S., 1971, Mutanti dell'orologio di Drosophila melanogaster. Proc. Natl. Acad. Sci., 68: 2112-2116.
- Konopka, R. J., Kyriacou, C. P. & Hall, J. C., 1996, Analisi mosaico nel Drosophila CNS dei ritmi del canto di corteggiamento circadiano influenzati da una mutazione dell'orologio periodico. J. Neurogenet., 11: 117-140.
- Kuhn, G. C. S., Diniz, N. M., Alves, M. A. R. & Sene, F. M., 1995, Costituzione cariotipica di specie del sottogruppo fasciola del gruppo repleta genere Drosophila. Rev. Bras. Gen., 18 supplemento: 282.
- Kyriacou, C. P. & Hall, J. C., 1980, Mutazioni del ritmo circadiano in Drosophila melanogaster influenzano le fluttuazioni a breve termine nel canto di corteggiamento del maschio. Proc. Natl. Acad. Sci., 77: 6729-6733.
- Manfrin, M. H., Prado, P. R. R. & SENE, F. M., 1997, Analisi dei componenti sonori del corteggiamento sessuale di due sottospecie di Drosophila mercatorum (Diptera, Drosophilidae). Rev. Brasil. Biol., 57: 349-355.
- Neems, R. M., Dooher, K., Butlin, R. K. & Shorrocks, B., 1997, Differenze nel canto di corteggiamento maschile tra le specie del gruppo quinaria di Drosophila. Journal of Insect Behavior, 10: 237-246.
- Noor, M. A. & Aquadro, C. F., 1998, Canti di corteggiamento di Drosophila pseudoobscura e D. persimilis: analisi della variazione. Anim. Behav., 56: 115-125.
- Ritchie, M. G. & Gleason, J. M., 1995, Evoluzione rapida del modello di canto di corteggiamento in specie di Drosophila willistoni. J. Evol. Biol., 8: 463-479.
- Ritchie, M. G. & Kyriacou, C. P., 1996, Selezione artificiale per un segnale di corteggiamento in Drosophila melanogaster. Anim. Behav., 52: 603-611.
- Ritchie, M. G., Yate, V. H. & Kyriacou, C. P., 1994, Variabilità genetica dell'intervallo interpulsato del canto di corteggiamento tra alcune popolazioni europee di Drosophila melanogaster. Eredità, 72: 459-464.
- Schilcher, F. V., 1976, Il ruolo degli stimoli uditivi nel corteggiamento di Drosophila melanogaster. Anim. Behav., 24: 18-26.
- Sene, F. M. & Manfrin, M. H., 1998, Una camera per registrare il canto di corteggiamento in Drosophila. D. I. S., 81: 203-204.
- Shorey, H. H., 1962, La natura del suono prodotto da Drosophila melanogaster durante il corteggiamento. Scienza, 137: 677-678.
- Tidon-Sklorz, R. & Sene, F. M., 1992, Distribuzione verticale e temporale delle specie di Drosophila (Diptera, Drosophilidae) in un'area boschiva nello stato di São Paulo, Brasile. Rev. Brasil. Biol., 52: 311-317.
- Tomaru, M. & Oguma, Y., 1994, Differenze nel canto di corteggiamento nelle specie del complesso Drosophila auraria. Anim. Behav., 47: 133-140.
- Vanzolini, P. E., 1981, Un approccio quasi-storico alla storia naturale della differenziazione dei rettili in isolate geografiche tropicali. Papeis Avulsos Zool., 34: 189.
- Vilela, C. R., 1983, Una revisione del gruppo di specie Drosophila repleta (Diptera, Drosophilidae). Revta. Bras. Ent., 27: 1-114.
- Vilela, C. R., Pereira, M. A. Q. R. & Sene, F. M., 1983, Dati preliminari sulla distribuzione geografica delle specie di Drosophila all'interno dei domini morfoclimatici del Brasile. II. Il gruppo repleta. Cienc. Cult., 35: 66-70.
- Wasserman, M., 1962, Studi citologici del gruppo repleta del genere Drosophila. VI. Il sottogruppo fasciola. Univ. Texas Public., 6205: 119-134.
- Wasserman, M., 1982, Evoluzione del gruppo repleta. In: M. Ashburner, H. L. Carson & J. N. Thompson (eds.), La genetica e la biologia di Drosophila, vol. 3b. Academic Press, London, pp. 61-139.
- Wasserman, M., 1992, Evoluzione citologica del gruppo di specie Drosophila repleta: VI. Il sottogruppo fasciola. In: Studi in Genetica II. Rapporti di ricerca sulla genetica di Drosophila, tassonomia ed evoluzione. CRC Press, Inc., M. R. Università del Texas, Austin.
- Welbergen, P., Van Duken, F. R., Scharloo, W. & Köhler, W., 1992, La base genetica dell'isolamento sessuale tra Drosophila melanogaster e D. simulans. Evoluzione, 46: 1385-1398.
- Wheeler, D. A., Fields, W. L. & Hall, J. C., 1988, Analisi spettrale dei canti di corteggiamento di Drosophila: D. melanogaster, D. simulans, e il loro ibrido interspecifico. Behav. Genet., 18: 675-703.