La comunità di Drosophila nelle vegetazioni xerofitiche del bacino superiore del fiume Paraná-Paraguay
Traduzione automatica
L'articolo originale è scritto in lingua EN (link per leggerlo) .
Abstract
Il genere Drosophila è il più abbondante nella famiglia Drosophilidae. Alcune specie sono endemiche di determinate regioni e altre sono cosmopolite. In Brasile, ci sono diversi ecosistemi da esplorare riguardo alla composizione e agli aspetti ecologici di Drosophila. Le aree xerofitiche sono un esempio. Possono essere trovate nel Sud e nel Sud-est del Brasile come isole, a causa dei cambiamenti nei cicli paleoclimatici. L'obiettivo del presente lavoro è fornire informazioni sulla composizione della comunità di Drosophila in otto aree xerofitiche (dove si trova il cactus Cereus hildmaniannus) nel Sud e nel Sud-est del Brasile. Questo lavoro rappresenta un passo importante nello studio degli aspetti ecologici quantitativi della comunità di Drosophila nelle aree xerofitiche che formano isole continentali nella regione neotropicale. La composizione della comunità di Drosophila trovata è compatibile con lavori precedenti in diversi aspetti. Gli indici ecologici hanno mostrato una possibile associazione positiva tra diversità e grado di preservazione delle aree studiate. Le comunità dello stato di São Paulo hanno presentato la massima somiglianza tra i siti testati, sebbene non sia stata trovata alcuna correlazione statistica significativa tra l'indice di Jaccard e la distanza geografica utilizzando il test di Mantel.
Introduzione
Sebbene nel XVIII secolo Scopoli, Linné e Fabricius siano stati i primi a descrivere le specie oggi allocate alla famiglia Drosophilidae, Duda (1924a, 1924b, 1925) può essere considerato il pioniere nella sistematica di questo gruppo. Inoltre, Sturtevant, noto per il suo lavoro classico (1921, 1939, 1942), è uno dei ricercatori più importanti all'interno di questa famiglia. I primi dati sulla comunità brasiliana di Drosophila furono pubblicati da Duda (1925). Nella regione sud-est del Brasile, Dobzhansky & Pavan (1943) furono pionieri e descrissero diverse nuove specie. Dopo il 1940, furono pubblicati molti lavori sulla fauna di Drosophila brasiliana (Pavan & Cunha, 1947; Dobzhansky & Pavan, 1950; Pavan, 1950; 1959) inclusa la prima lista di specie di Drosophila brasiliane (Mourão et al., 1965). Successivamente, altri autori analizzarono la fauna di Drosophila nei domini morfoclimatici del Brasile nel tentativo di comprendere meglio la distribuzione geografica delle specie più comuni (Sene et al., 1980; Vilela et al., 1983; Val et al., 1981).
La famiglia Drosophilidae è composta da 65 generi e più di 3.500 specie descritte che si trovano in un numero di ecosistemi in tutto il mondo (Bächli, 1998). La maggior parte dei generi si trova nelle regioni tropicali. Il genere Drosophila è il più abbondante e comprende circa il 53% delle specie totali. Alcune di esse sono endemiche di determinate regioni e altre sono cosmopolite, disperse principalmente in associazione con l'attività umana. Grimaldi (1990) ha esaminato la classificazione della famiglia Drosophilidae basandosi su un'analisi cladistica utilizzando la massima parsimonia. Questo autore ha studiato 217 caratteri morfologici degli adulti e 120 specie rappresentative della famiglia e ha concluso che costituiscono un gruppo monofiletico. Il genere Drosophila probabilmente è originato nei tropici (Stalker, 1976) e diversi membri di esso hanno ampliato la loro distribuzione nelle zone temperate, dove è stata selezionata la capacità di utilizzare risorse in condizioni ambientali estreme. Questo processo potrebbe essersi ripetuto più volte durante l'evoluzione del genere.
Oggi, la maggior parte della ricerca che utilizza le specie di Drosophila viene condotta nei campi della Biologia Molecolare e della Genetica, ma solo all'inizio del ventesimo secolo Drosophila melanogaster è stata riconosciuta come utile per le analisi genetiche (Castle, 1906). Nonostante il gran numero di studi che utilizzano Drosophila come organismo modello in questi campi, le specie brasiliane richiedono ancora ulteriori studi riguardo a molti aspetti di ecologia, sistematica, genetica ed evoluzione.
Alcuni studi ecologici con Drosophila sono stati condotti, specialmente riguardo all'attività delle mosche durante il giorno (Klaczko et al., 1983; Medeiros, 2000), il periodo appropriato della giornata per il campionamento (Pavan et al., 1950; Belo & Oliveira-Filho, 1978), e la distribuzione spaziale delle specie (orizzontale: Dobzhanzky & Pavan, 1950; Burla et al., 1950; Pavan, 1959; Medeiros, 2000; e verticale: Sene et al., 1981; Kratz et al., 1982 e Tidon-Sklorz & Sene, 1992). Tuttavia, il confronto tra aree ha ricevuto poca attenzione (Medeiros & Klaczko, 2004, per esempio) e rimane ancora un'area di studio aperta per le specie brasiliane di Drosophila.
Tenendo conto di ciò, ci sono diversi ecosistemi da esplorare in Brasile riguardo alla composizione e agli aspetti ecologici di Drosophila. Le aree xerofitiche, determinate dalla presenza di vegetazione di tipo secco con l'occorrenza di diversi tipi di cactus, sono un esempio. Attualmente, così come le aree centrali di occorrenza dei cactus, come la Caatinga brasiliana e il Chaco argentino, queste piante possono essere trovate nel Sud, nel Sud-est e nelle regioni Centrali del Brasile su isole, risultato dei cambiamenti paleoclimatici ciclici. Questi cicli alternavano tra freddo/secco e caldo/umido. Nel primo caso, le aree xerofitiche si espandevano dalle aree centrali sul continente e nel secondo, si ritraevano formando rifugi (isole) tra di esse (Bigarella et al., 1975; Ab’Saber, 1977; Vanzolini, 1981). Pertanto, i rifugi sono importanti per la biodiversità, per gli studi ecologici ed evolutivi di diversi gruppi, comprese le specie di Drosophila che si riproducono specificamente nel tessuto di cactus in decomposizione, come fanno le mosche del gruppo D. repleta.
L'obiettivo del presente lavoro è fornire informazioni sulla composizione della comunità di Drosophila in otto aree xerofitiche (dove si trova il cactus Cereus hildmaniannus ) nelle regioni sud e sud-est del Brasile. Questo fornirà informazioni preziose per ulteriori studi riguardanti l'ecologia, la genetica e l'evoluzione delle popolazioni di Drosophila geograficamente isolate.
Materiali e metodi
I luoghi e le date delle raccolte sono riassunti nella Tabella 1. I campioni sono stati raccolti in boschi composti da vegetazione xerofitica in foreste mesofitiche nei vallate della parte superiore del bacino idrografico del fiume Paraná-Paraguay. Queste aree si trovano in diversi stadi di perturbazione, che vanno dall'uso per l'allevamento del bestiame accanto a una strada (Itatiba - SP e Sengés - PR) a un'area di conservazione del Parco Statale (Canyon Guartelá - PR). Tutte le raccolte sono state effettuate alla fine del periodo umido, che corrisponde a febbraio-maggio in due anni diversi (1999 e 2000).
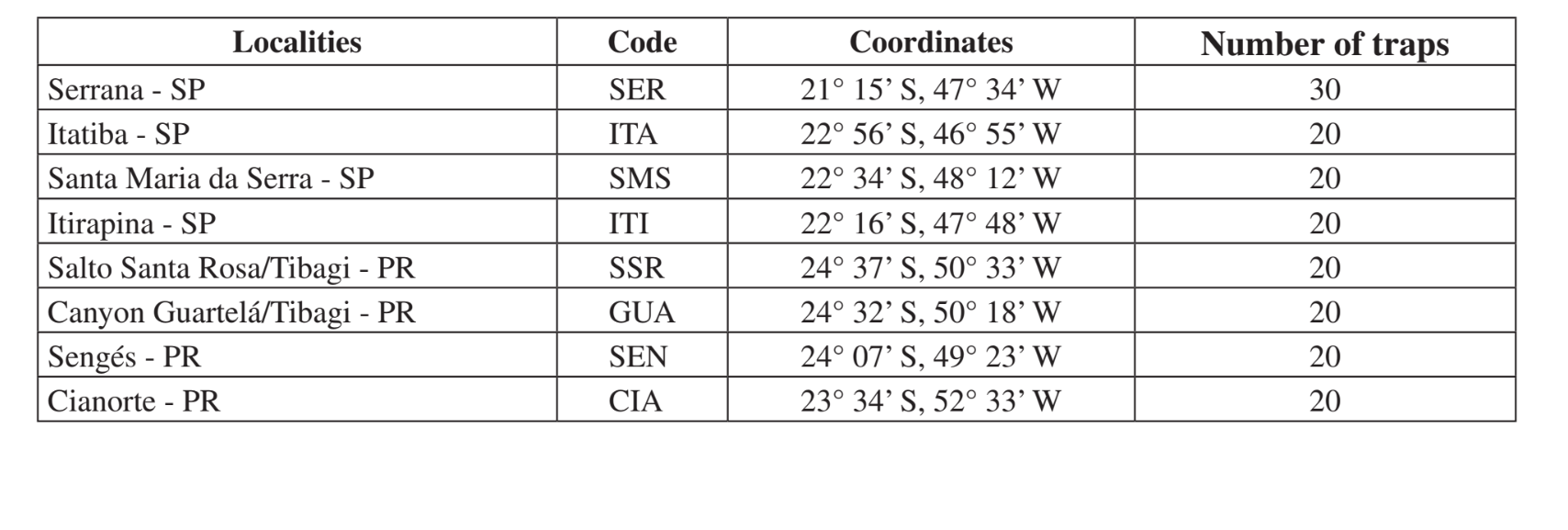
Collezioni
Per tre giorni, le mosche adulte sono state attratte da banana e arancia fermentate in trappole aperte, disperse non più vicine di 15 m l'una dall'altra, appese a rami a ± 1,5 m di altezza dal suolo. Dopo questo periodo, le mosche sono state catturate con reti entomologiche, messe in barattoli di vetro con terreno di coltura e portate in laboratorio vive, dove sono state identificate.
È difficile identificare i membri dei gruppi Drosophila repleta e D. willistoni a un certo grado. Le femmine del primo gruppo sono considerate non identificate se la caratteristica diagnostica è l'aedeago maschile. Per quanto riguarda il gruppo Drosophila willistoni, l'identificazione utilizzando tratti genetici è più affidabile ma non è stata effettuata. Pertanto, le specie crittiche sono state raggruppate come “repleta gruppo” e “D. willistoni gruppo”, rispettivamente. Le specie non identificate simili al “repleta gruppo” non sono state considerate nelle analisi.
Analisi dei dati
Per verificare qualitativamente l'occorrenza delle specie, è stato utilizzato il Metodo di Costanza dell'Ocorrenza (Dajoz, 1983). Il valore di costanza (c) è stato ottenuto dividendo il numero di raccolte in cui è comparsa una specie per il numero totale di raccolte, e poi moltiplicando quel risultato per 100. Le specie con indice c ≥ 50 sono state considerate costanti. Le specie accessorie erano quelle con 25 ≤ c < 50. Le specie accidentali avevano c < 25. Le specie che si sono verificate in un'unica area sono state considerate esclusive.
Le comunità di Drosophila sono state analizzate utilizzando diversi indici ecologici: Berger-Parker (1/d), Shannon-Wienner (H’), Simpson (D), Margalef (DMg), Menhinick (DMn) e Jaccard (J). È stata utilizzata un'ANOVA non parametrica, Kruskal-Wallis, per determinare se ci fossero differenze nella abbondanza mediana tra le comunità. Successivamente, è stata effettuata una comparazione multipla tra i campioni (simile al test di Tukey) per determinare se si verificassero differenze significative tra i campioni (Zar, 1999).
Per verificare le relazioni tra le comunità, sono state eseguite due analisi di clustering:
1) utilizzando la presenza e l'assenza di specie; e 2) utilizzando il numero di esemplari raccolti per ogni specie in ciascuna area. I fenogrammi sono stati costruiti in Minitab® per Windows Release 10.1 selezionando la distanza euclidea e i metodi di collegamento singolo in entrambi gli approcci.
Per analizzare se esiste un'associazione tra le somiglianze delle comunità e le distanze geografiche, è stato effettuato un test di Mantel utilizzando il software TFPGA (Miller, 1997) confrontando due matrici: indici di Jaccard e distanze geografiche tra coppie di comunità.
Risultati e discussione
In questo studio siamo stati in grado di identificare un totale di 10.464 mosche appartenenti a 4 sottogeneri, 10 gruppi e almeno 30 specie diverse di Drosophila (Tabella 2). Le specie costanti (c ≥ 50) rappresentavano circa il 63% del totale delle specie raccolte (19 su 30). Sette specie sono state considerate accessorie (~ 23%) e sono state trovate 4 specie accidentali (~ 13%). Tutti i sottogeneri hanno mostrato specie costanti, tranne il sottogenere Dorsilopha che ha mostrato solo una specie accidentale (Drosophila busckii).
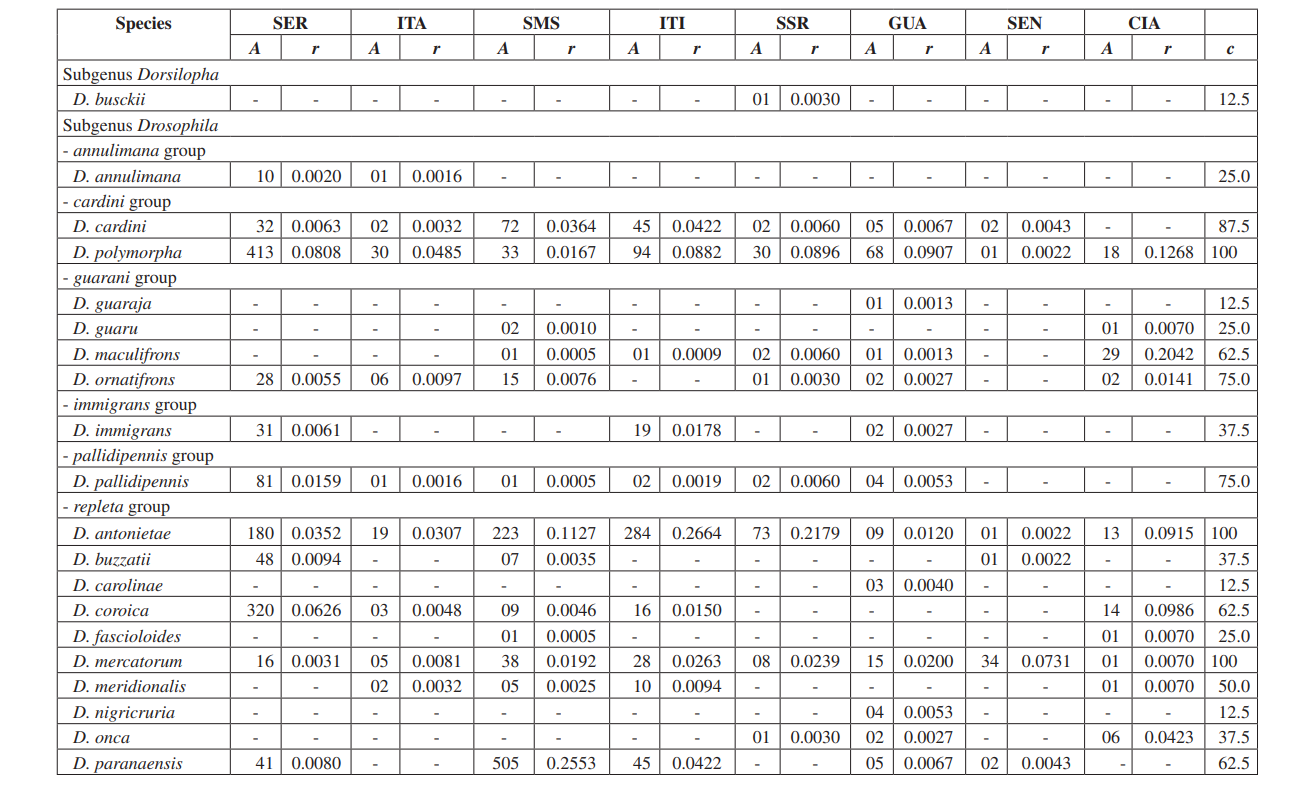
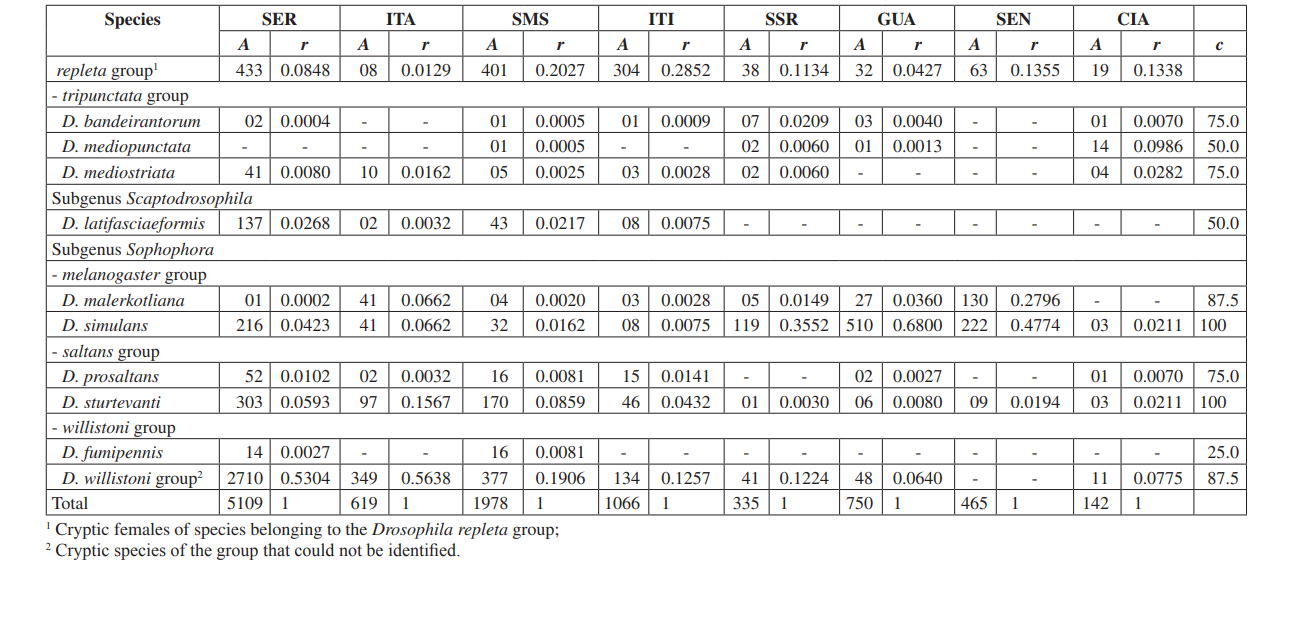
Cinque specie (Drosophila polymorpha, D. antonietae, D. mercatorum, D. simulans e D. sturtevanti) sono state considerate abbondanti e sono state raccolte in tutte le 8 aree (Serrana - SP, Itatiba - SP, Santa Maria da Serra - SP, Itirapina - SP, Salto Santa Rosa - PR, Canyon Guartelá - PR, Sengés - PR e Cianorte - PR). Questo risultato è compatibile con precedenti indagini su Drosophila.
Drosophila polymorpha è stata raccolta dal Guatemala al Brasile e in diversi tipi di ambienti: foreste, cerrado, restinga, e anche associata alla presenza umana. È una specie relativamente abbondante in diversi domini morfoclimatici, tranne che nella caatinga (Sene et al., 1980). Drosophila antonietae era prevista in tutte le aree, poiché è una specie cactofila che si trova nelle regioni sud e sud-est del Brasile, e a nord del confine orientale del Chaco argentino. Questa specie si trova sempre associata al cactus colonnare Cereus hildmaniannus nella vegetazione xerofitica delle foreste a galleria e mesofitiche nei vallate del bacino del fiume Paraná-Uruguai (Tidon-Sklorz & Sene, 2001). Questo modello di distribuzione che isola Drosophila antonietae da altre specie del gruppo D. buzzatii coincide con modelli definiti di altri ditteri e anche con regioni di endemismo, suggerendo eventi vicarianti come barriere zoogeografiche stabilite in epoche geologiche precedenti o eventi climatici che hanno impedito il flusso genico consentendo l'accumulo di differenze tra le popolazioni (Amorin & Pires, 1996). Questo modello di distribuzione è osservato anche riguardo alla distribuzione delle piante ristrette alle regioni xerofitiche della caatinga e del chaco (Duvernell & Eanes, 2000).
Drosophila mercatorum è abbastanza comune negli ambienti naturali in Sud America, specialmente in aree aperte (Sene et al., 1981; Vilela et al., 1983).
Drosophila simulans è una specie introdotta che è stata raccolta ad alta frequenza in diversi luoghi (Dobzhansky & Pavan, 1950; Pavan, 1959; Sene et al., 1980; Tidon-Sklorz & Sene, 1992), anche in aree aperte (Perondini et al., 1979).
Drosophila sturtevanti è una specie ampiamente distribuita in America Centrale e Meridionale che si verifica in diversi domini morfoclimatici (Tidon-Sklorz & Sene, 1999).
Specie esclusive sono apparse a Salto Santa Rosa - PR (Drosophila busckii) e Canyon Guartelá - PR (Drosophila guaraja, D. carolinae e D. nigricruria). Drosophila busckii non era attesa a essere raccolta a Salto Santa Rosa - PR poiché è una specie introdotta in Brasile ed è frequentemente associata a ambienti modificati dall'uomo (Tidon-Sklorz & Sene, 1999). Tuttavia, è stata raccolta in ambienti naturali principalmente in vegetazioni aperte secche (Bizzo & Sene, 1982; Tidon-Sklorz & Sene, 1992). Tra le specie esclusive raccolte nel Canyon Guartelá - PR, Drosophila guaraja è stata registrata in Brasile solo a Boracéia - SP (Val & Kaneshiro, 1988). Questo è, quindi, il primo reperto al di fuori di questa località in Brasile. Drosophila carolinae è stata raccolta solo nelle foreste pluviali in Brasile (Vilela, 1983) e per questo motivo non ci si aspettava di trovarla in aree di vegetazione xerofitica. Tuttavia, il Canyon Guartelá - PR è una delle aree più preservate indagate e i cactus sono distribuiti sotto una regione boschiva all'interno del Parco Statale, un fatto che potrebbe spiegare la sua presenza. Drosophila nigricruria è l'unica specie esclusiva che ha una vasta distribuzione geografica. Tuttavia, è stata raccolta solo in quest'area in questo studio. Medeiros & Klaczko (2004) hanno anche raccolto questa specie nella fattoria Barreiro Rico (tra 48° 05’ 11’’ W – 22° 41’ 15’’ S e 48° 04’ 52’’ W – 22° 41’ 26’’ S), un plateau interno che ha una foresta semi-decidua e una delle tre foreste rimanenti nello Stato di São Paulo dove sono state effettuate raccolte da questi autori.
Il Drosophila subgenere ha mostrato la massima ricchezza (7 gruppi e 22 specie), con 7 specie appartenenti al gruppo Drosophila repleta . La seconda massima ricchezza è stata trovata nel subgenere Sophophora (3 gruppi e 6 specie). Questa distribuzione della ricchezza tra i subgeneri è stata osservata in altri studi (Sene et al., 1981; Tidon-Sklorz & Sene, 1995; Tidon-Sklorz & Sene, 1999; Medeiros & Klaczko, 2004).
La specie di drosofila introdotta, Zaprionus indianus (Vilela, 1999), è stata trovata in tutte le collezioni (1.459 esemplari a Serrana; 317 a Itatiba; 409 a Santa Maria da Serra; 302 a Itirapina; 42 a Salto Santa Rosa; 117 a Guartelá; 598 a Sengés; e 8 a Cianorte). Gli eventi associati alle invasioni biologiche sono divisi in tre categorie: arrivo, stabilizzazione e integrazione (Vermeij, 1996). Secondo Moraes et al. (2000), Zaprionus indianus ha attraversato tutte e tre e probabilmente sta competendo sfavorevolmente con altre specie simili nel sud-est del Brasile, poiché è stata osservata una diminuzione della frequenza relativa. L'introduzione di una specie esotica di Drosophila in Brasile non è un evento raro. Nel 1976, Val & Sene (1980) raccolsero un gran numero di Drosophila malerkotliana nel Brasile nordorientale, una specie che non era stata registrata in ampie collezioni precedenti. Attualmente, nei nostri sondaggi, Drosophila malerkotliana viene raccolta quasi in tutto il Brasile, sebbene sempre a una bassa frequenza relativa.
Ventitré specie diverse sono state raccolte a Santa Maria da Serra - SP, 20 specie a Serrana - SP e nel Canyon Guartelá - PR, 18 a Itirapina - SP, 17 a Cianorte - PR, 16 a Itatiba - SP e Salto Santa Rosa - PR e 9 a Sengés - PR (Tabella 3). Medeiros & Klaczko (2004) hanno descritto l'ultimo inventario della fauna di Drosophila in Brasile, indagato in tre foreste residue nello Stato di São Paulo. Questi autori hanno ottenuto 57, 76 e 90 specie diverse nelle tre aree studiate. Questi numeri di specie sono superiori a quelli osservati qui, forse a causa di differenze nel metodo di raccolta o semplicemente perché le aree indagate sono distinte nella loro costituzione vegetale (foresta e vegetazione xerofitica). La massima abbondanza totale è stata rilevata a Serrana - SP (4676 esemplari) e la minima a Cianorte - PR (123 esemplari) (Tabella 3).
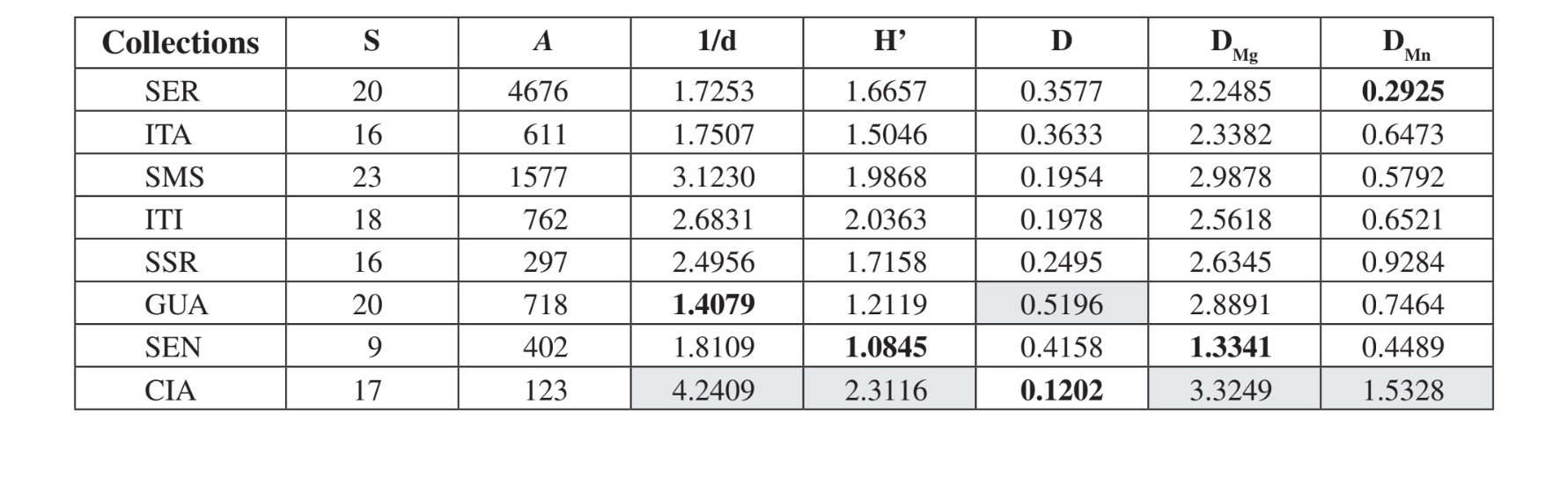
Secondo gli indici ecologici (Tabella 3), Cianorte - PR ha presentato valori più elevati in quasi tutti gli indici (1/d = 4.2409; H’ = 2.3116; DMg = 3.3249; DMn = 1.5328), tranne che per Simpson (D) che ha mostrato Canyon Guartelá - PR come l'area più
diversa (D = 0.5196). Canyon Guartelá - PR ha anche mostrato uno dei valori più alti in altri due casi (DMg = 2.8891; DMn = 0.7464). Sengés - PR, Itatiba - SP e Serrana - SP hanno mostrato i valori più bassi per la maggior parte degli indici. Santa Maria da Serra - SP, Itirapina - SP e Salto Santa Rosa - PR hanno mostrato valori intermedi per la maggior parte degli indici. Queste osservazioni sono compatibili con la valutazione superficiale della degradazione in queste aree. Cianorte - PR si trova vicino a una strada, ma su una ripida sponda del fiume Ligeiro, con accesso difficile per la degradazione umana. Canyon Guartelá - PR è un'area boschiva all'interno del Parco Statale Canyon Guartelá dello stato del Paraná, come menzionato in precedenza. Pertanto, è un'area altamente preservata. Sengés - PR è l'area più degradata, vicino a una strada, sede di allevamento di bestiame e sotto l'alta influenza dell'attività umana. Serrana - SP è un'area preservata sulla cima di una collina che è difficile da raggiungere. I valori di diversità più bassi trovati per quest'area devono essere dovuti al numero maggiore di trappole utilizzate in questa raccolta (Tabella 1). Le altre aree ci si aspettava avessero valori intermedi di diversità, poiché sono piccole aree di conservazione all'interno di fattorie.
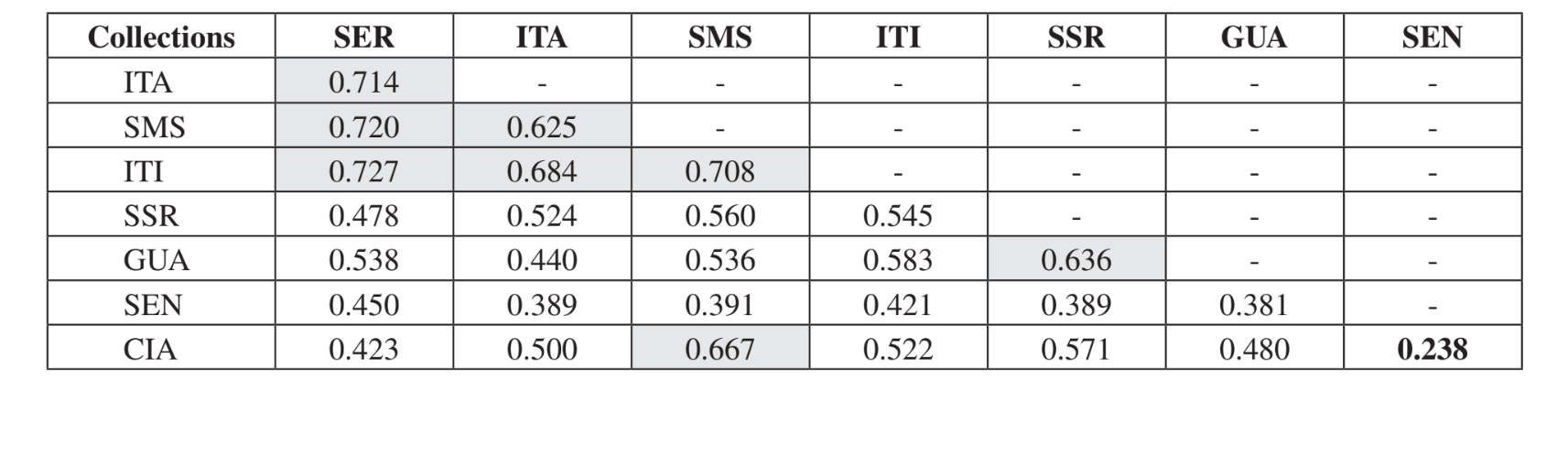
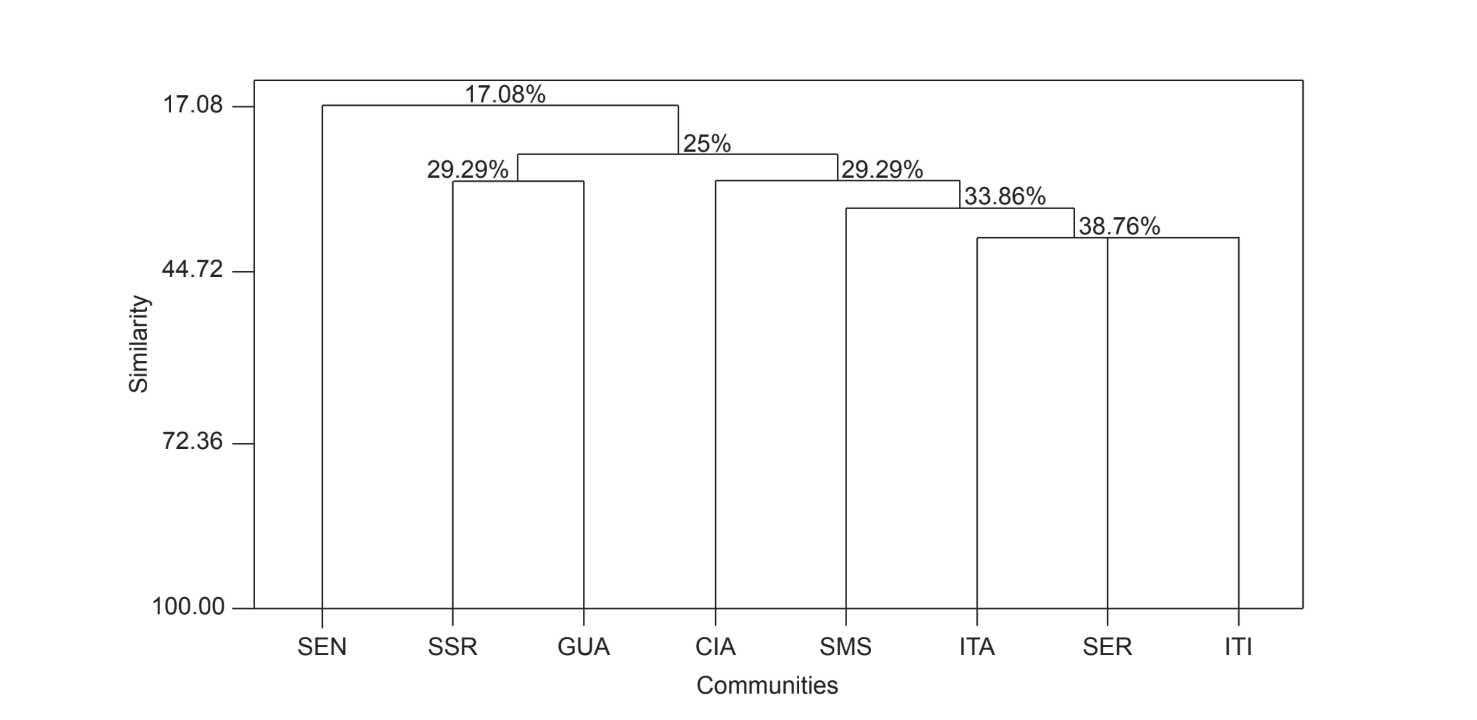
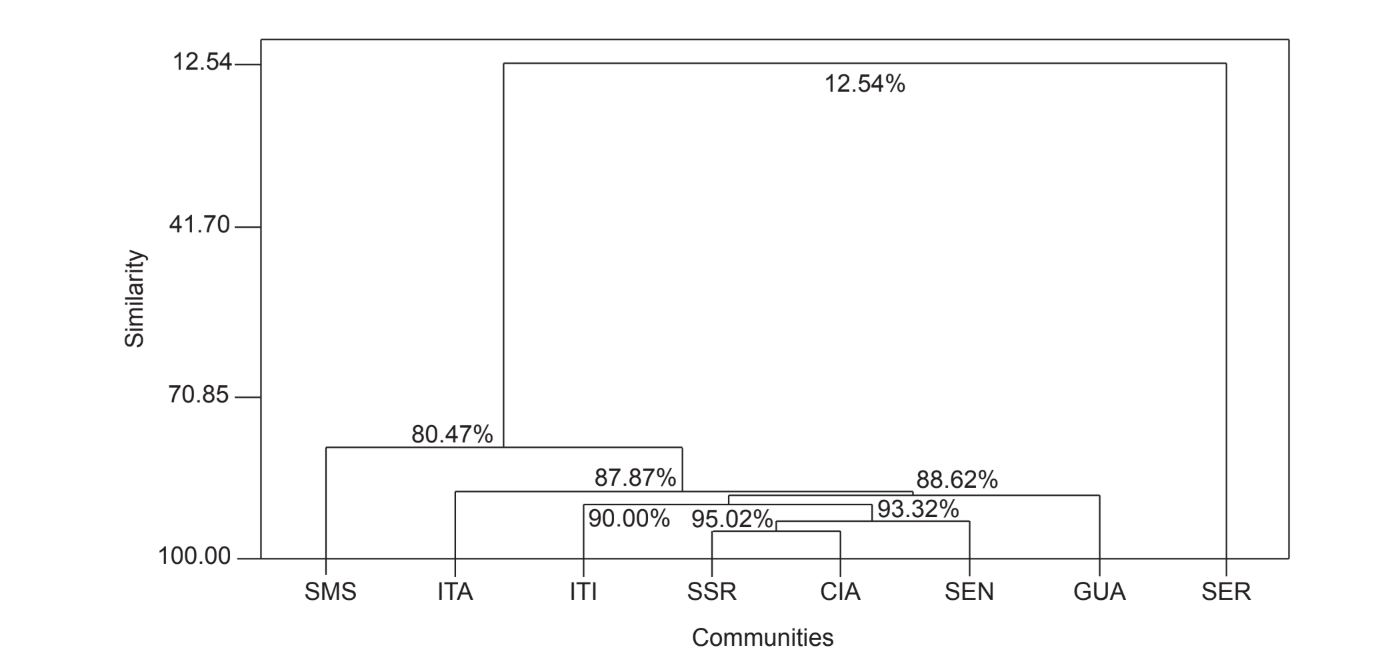
Il confronto a coppie delle comunità utilizzando l'indice di Jaccard (Tabella 4) ha mostrato che le comunità dello stato di São Paulo (Serrana - SP, Itatiba - SP, Santa Maria da Serra - SP e Itirapina - SP) avevano la massima somiglianza tra tutti i confronti (J > 0.62). Lo stesso modello non è stato osservato tra le comunità dello stato del Paraná e anche tra i due stati, ad eccezione dell'indice ottenuto tra Santa Maria da Serra - SP e Cianorte - PR (J = 0.667), e tra Salto Santa Rosa - PR e Canyon Guartelá - PR (J = 0.636). Le analisi di clustering utilizzando i dati di presenza e assenza delle specie (Fig. 1), nonostante un basso livello di somiglianza (33.86%), hanno comunque raggruppato tutte le comunità dello stato di São Paulo insieme. La comunità più divergente è stata Sengés - PR. La Fig. 2 illustra il raggruppamento delle comunità utilizzando i dati di abbondanza assoluta (A). Sono stati trovati livelli di somiglianza più elevati, ma non è stata riscontrata una chiara associazione tra le comunità. In questa figura, Serrana - SP era la comunità più diversa.
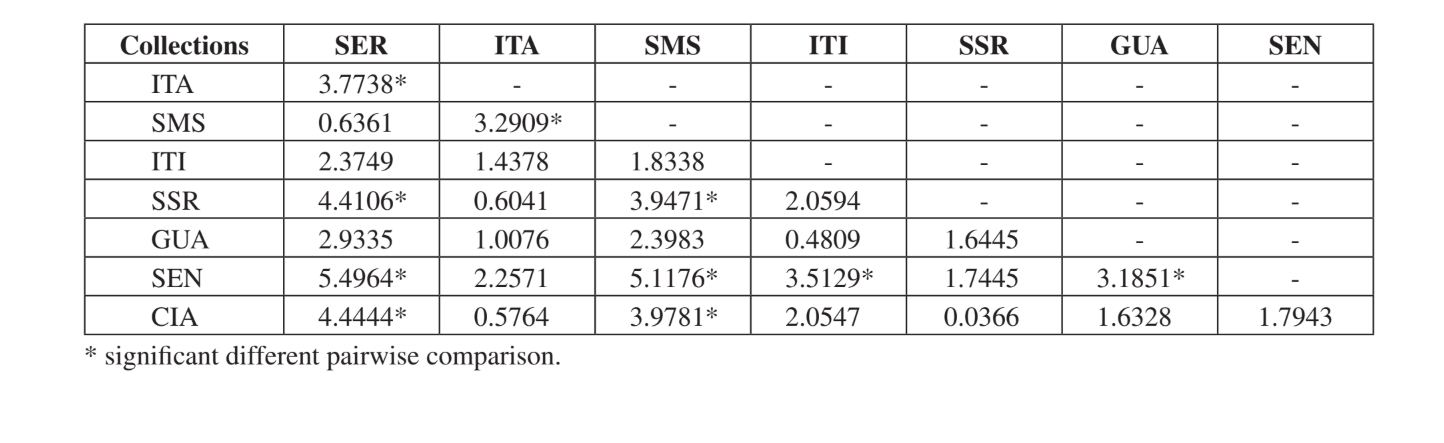
Il test di Kruskal-Wallis, corretto per i legami, ha mostrato che le comunità sono significativamente diverse quando è stata effettuata una comparazione generale (H = 24.24; d.f. = 7; p = 0.001). I confronti multipli a coppie delle comunità sono presentati nella Tabella 5. Le comunità di Serrana - SP, Santa Maria da Serra – SP e Sengés - PR erano le più distinte quando confrontate individualmente con le altre (4 confronti significativamente diversi per ciascuna). Itirapina - SP e Canyon Guartelá - PR erano le comunità più simili quando confrontate con ciascuna comunità campionata (solo una differenza significativa per ciascuna). Tutti questi confronti (indice di Jaccard, analisi di cluster e confronti multipli a coppie) hanno mostrato che una maggiore somiglianza si è verificata tra le comunità dello Stato di São Paulo, sebbene non sia stata trovata alcuna correlazione positiva significativa nel test di Mantel che confronta l'indice di Jaccard e le distanze geografiche (r = – 0.3235; p = 0.0839).
Il sondaggio sulla composizione della comunità è un passo importante per diversi studi in un'ampia gamma di campi, come genetica, ecologia ed evoluzione. Questo lavoro aggiunge nuove conoscenze riguardo alla composizione e alla diversità della comunità di Drosophila in aree xerofitiche che formano isole continentali nella regione neotropicale. Questo approccio è importante per ulteriori studi in qualsiasi campo che consideri Drosophila come modello.
Autori: Mateus, R. P., Buschini, M. L. T. e Sene, F. M
Riferimenti:
- Ab’saber, A. N., 1977, Spazi occupati dall'espansione dei climi secchi dell'America del Sud, in occasione dei periodi glaciali quaternari. Paleoclimas, 3: 1-19.
- Amorin, D. S. & Pires, M. R. S., 1996, Biogeografia neotropicale e metodo per la stima massima della biodiversità. In: C. E. M. Bicudo & N. A. Menezes (eds.), Biodiversità in Brasile, un primo approccio, CNPq, São Paulo.
- Bächli, G., 1998, Famiglia Drosophilidae. In: L. Papp & B. Darvas (eds.), Contributi a un Manuale di Diptera Paleartica. III. Brachicera Superiore, Science Herald, Budapest.
- Belo, M. & Oliveira-Filho, J. J., 1978, Specie domestiche di Drosophila. V. Influenze di fattori ambientali sul numero di individui catturati. Rev. Bras. Biol., 36: 903-909.
- Bigarella, J. J., Andrade-Lima, D. & Riehs, P. J., 1975, Considerazioni riguardo ai cambiamenti paleoclimatici nella distribuzione di alcune specie vegetali e animali in Brasile. An. Acad. Brasil. Ciênc., 41: 411-464.
- Bizzo, N. M. V. & Sene, F. M., 1982, Studi sulle popolazioni naturali di Drosophila da Peruíbe (SP), Brasile (Diptera, Drosophilidae). Rev. Bras. Biol., 42: 539-544.
- Burla, H., da Cunha, A. B., Cavalcanti, A. G. L., Pavan, C. & Dobzhansky, T., 1950, Densità di popolazione e tassi di dispersione in Drosophila willistoni brasiliana. Ecology, 31: 393-404.
- Castle, W. E., 1906, Inbreeding, incrocio e sterilità in Drosophila. Science, 23: 153.
- Dajoz, R., 1983, Ecologia Generale. 471p. Editora Vozes, Petrópolis.
- Dobzhansky, T. & Pavan, C., 1943, Studi sulle specie brasiliane di Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 36 (Biol. Geral, 4): 1-72.
- Dobzhansky, T. & Pavan, C., 1950, Variazioni locali e stagionali nelle frequenze relative delle specie di Drosophila in Brasile. J. Anim. Ecol., 19: 1-14.
- Duda, O., 1924a, Contributo alla sistematica dei Drosophilidi con particolare attenzione alle specie paleartiche e orientali (Diptera). Arch. Naturgesch., 90: 172-234.
- Duda, O., 1924b, Revisione delle specie europee del genere Drosophila Fallén (Diptera). Ent. Medd., 14: 246-313.
- Duda, O., 1925, I Drosophilidi costaricensi del Museo Nazionale Ungherese di Budapest. Annls hist.- nat. Mus. natn. hung., 22: 149-229.
- Duvernell, D. D. & Eanes, W. F., 2000, Genetica di popolazione molecolare contrastante di quattro esochinasi in Drosophila melanogaster, D. simulans e D. yakuba. Genetics, 156: 1191-1201.
- Grimaldi, D. A., 1990, Una classificazione filogenetica rivista dei generi nella Drosophilidae (Diptera). Bull. Am. Mus. nat. Hist., 197: 1-139.
- Klaczko, L. B., Powell, J. R. & Taylor, C. E., 1983, Drosophila esche e lieviti: specie attratte. Oecologia, 59: 411-413.
- Kratz, F. L., Pinto, L. G., Brandão, D. & Faria, L. G., 1982, Altezza di volo e il modello di distribuzione spaziale in Drosophila. Ciênc. Cult., 34: 203-209.
- Medeiros, H. F., 2000, Assembléias di Specie di Drosophila (Diptera; Drosophilidae) e Effetti di Corsi d'Acqua sulle loro Distribuzioni in due Matas di São Paulo, 98p. Dissertazione (Mestrado) – Instituto di Biologia IB/UNICAMP, Campinas.
- Medeiros, H. F. & Klaczko, L. B., 2004, Quante specie di Drosophila (Diptera, Drosophilidae) rimangono da descrivere nelle foreste di São Paulo, Brasile? Elenchi di specie di tre resti forestali. Biota Neotropica, 4: 1-12.
- Miller, M. P., 1997, Strumenti per analisi genetiche di popolazione - TFPGA - 1.3: Un programma Windows per l'analisi di dati genetici di allozimi e di popolazione molecolare.Software distribuito dall'autore.
- Moraes, E. M., Mateus, R. P. & Sene, F. M., 2000, Occorrenza e fluttuazione nella dimensione della popolazione di una specie di Drosophilidae recentemente introdotta in Brasile. In: ASTRAZIONI DEL XXI CONGRESSO INTERNAZIONALE DI ENTOMOLOGIA, LIBRO I. Atti..., 271p.
- Mourão, C. A., Gallo, A. J. & Bicudo, H. E. M. C., 1965, Sulla sistematica di Drosophila in Brasile, con la descrizione di D. mendeli sp.n. e “Relazione di Specie Brasiliane del Genere Drosophila”. Ciênc. Cult., 17: 577-585.
- Pavan, C., 1950, Specie Brasiliane di Drosophila. II. Bolm. Fac. Filos. Ciênc. S. Paulo, 111 (Biol. Geral, 8): 1-37.
- Pavan, C., 1959, Relazioni tra popolazioni naturali di Drosophila e l'ambiente. Bolm. Fac. Filos. Ciênc. S. Paulo, 221 (Biol. Geral, 11): 1-81.
- Pavan, C. & da Cunha, A. B., 1947, Specie Brasiliane di Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 86 (Biol. Geral, 7): 3-47.
- Pavan, C., Dobzhansky, T. & Burla, H., 1950, Comportamento diurno di alcune specie neotropicali di Drosophila. Ecology, 31: 36-43.
- Perondini, A. L. P., Sene, F. M. & Mori, L., 1979, Il modello e il polimorfismo di alcune esterasi di Drosophila simulans in Brasile. Egypt. J. Genet. Cytol., 8: 263-268.
- Sene, F. M., Val, F. C., Vilela, C. R. & Pereira, M. A. Q. R., 1980, Dati preliminari sulla distribuzione geografica delle specie di Drosophila all'interno dei domini morfoclimatici del Brasile. Pap. Avul. Dep. Zool. Sec. Agric., 33: 315-326.
- Sene, F. M., Pereira, M. A. Q. R., Vilela, C. R. & Bizzo, N. M. V., 1981, Influenza di diversi modi di impostare esche per la raccolta di mosche di Drosophila in tre ambienti naturali. D. I. S., 56: 118-121..
- Stalker, H. D., 1976, Studi sui cromosomi in popolazioni selvatiche di D. melanogaster. Genetics, 82: 323-347.
- Sturtevant, A. H., 1921, Le specie nordamericane di Drosophila. Carnegie Inst. Wash. Publ., 301: 1-150.
- Sturtevant, A. H., 1939, Sulla suddivisione del genere Drosophila. Proc. Natl. Acad. Sci. USA, 3: 137-141.
- Sturtevant, A. H., 1942, La classificazione del genere Drosophila, con descrizioni di nove nuove specie. Univ. Texas Publ., 4213: 6-51.
- Tidon-Sklorz, R. & Sene, F. M., 1992, Distribuzione verticale e temporale delle specie di Drosophila (Diptera, Drosophilidae) in un'area boschiva nello stato di São Paulo, Brasile. Rev. Bras. Biol., 52: 311-317.
- Tidon-Sklorz, R. & Sene, F. M., 1995, Fauna di Drosophila (Diptera, Drosophilidae) nell'area settentrionale della “Cadeia do Espinhaço”, Stati di Minas Gerais e Bahia, Brasile: Aspetti biogeografici ed ecologici. Iheringia, Sér. Zool., 78: 85-94.
- Tidon-Sklorz, R. & Sene, F.M., 1999, Drosophila. In: C. R. F. Brandão & E. M. Cancello (eds), Invertebrati Terrestri. Vol.V. Biodiversità dello Stato di São Paulo. Sintesi della Conoscenza alla Fine del XX Secolo, FAPESP, São Paulo.
- Tidon-Sklorz, R. & Sene, F. M., 2001, Due nuove specie del set di fratelli di Drosophila serido (Diptera, Drosophilidae). Iheringia, Sér. Zool., 90: 141-146.
- Val, F. C. & Kaneshiro, K. Y., 1988, Drosophilidae (Diptera) dall'Estação Biológica de Boracéia, sulla catena costiera dello Stato di São Paulo, Brasile: Distribuzione geografica. In: P. E. Vanzolini & W. R. Heyer (eds.), Atti di un Workshop sui Modelli di Distribuzione Neotropicali, Academia Brasileira de Ciências, Rio de Janeiro.
- Val, F. C. & Sene, F. M., 1980, Una nuova specie di Drosophila introdotta in Brasile. Pap. Avul. Dep. Zool. Sec. Agric., 33: 293-298.
- Val, F. C., Vilela, C. R. & Marques, M. D., 1981, Drosophilidae della Regione Neotropicale. In: M. Ashburner, H. L. Carson & J. N. Thompson (eds.), La genetica e la biologia di Drosophila, Academic Press, Londra.
- Vanzolini, P. E., 1981, Un approccio quasi-storico alla storia naturale della differenziazione dei rettili in isolate geografiche tropicali. Pap. Avul. Dep. Zool. Sec. Agric., 34: 189-204
- Vermeij, G. J., 1996, Un'agenda per la biologia delle invasioni. Biol. Conserv., 78: 3-9.
- Vilela, C. R., 1983, Una revisione del gruppo di specie Drosophila repleta (Diptera, Drosophilidae). Rev. Bras. Entomol., 27: 114.
- Vilela, C. R., 1999, Zaprionus indianus Gupta, 1970, (Diptera, Drosophilidae) sta attualmente colonizzando la regione neotropicale? D. I. S., 82: 37-39.
- Vilela, C. R., Pereira, M. A. Q. R. & Sene, F. M., 1983, Dati preliminari sulla distribuzione geografica delle specie di Drosophila all'interno dei domini morfoclimatici del Brasile.II. Il gruppo repleta. Ciênc. Cult., 35: 66-70.
- Zar, J. H., 1999, Confronti Multipli. In: Analisi Biostatistica. 4a ed., Prentice Hall Inc., New Jersey.