
Somiglianza alloenzimatica e cromosomica in due specie di Drosophila
Traduzione automatica
L'articolo originale è scritto in lingua EN (link per leggerlo) .
Abstract
D. setosimentum e ochrobasis sono una coppia di specie molto vicine, parzialmente simpatiche, endemiche dell'isola di Hawaii. I maschi delle due specie differiscono in modo sorprendente nel motivo delle ali e ci sono differenze altitudinali e nei siti di riproduzione. Sono stati calcolati indici di somiglianza per entrambe le varianti cromosomiche (C) e allozimiche (A). All'interno delle popolazioni principali di ciascuna specie, entrambi i tipi di dati forniscono coefficienti superiori a 0,98. Il confronto interspecifico delle popolazioni principali mostra 0,66 (C) e 0,79 (A). Una popolazione isolata di ochrobasis dai Vulcani di Kohala (Ohu), quando confrontata con setosimentum, mostra 0,68 (C) e 0,98 (A). I cromosomi sono quindi molto più sensibili degli allozimi nel distinguere queste specie; lo stesso vale per D. silvestris e heteroneura provenienti dalle stesse foreste. Queste specie morfologicamente distinte, quando confrontate, mostrano 0,96 (A). Tutte e quattro le specie sembrano essere molto nuove in senso storico. In un'area, circa il 2% di D. setosimentum/ochrobasis catturati in natura sono ibridi interspecifici, sebbene campioni adeguati indichino che le popolazioni genetiche separate non si siano disintegrate. I nomi specifici dovrebbero essere mantenuti, ma le due entità sono forse meglio descritte come semispecie piuttosto avanzate in cui l'isolamento riproduttivo in natura è ora quasi completo.
Una rivoluzione è in corso nella sistematica contemporanea, specialmente a livello delle differenze tra specie (1). I dati sulla variabilità genetica all'interno e tra popolazioni strettamente correlate possono essere utilizzati per stabilire indici di somiglianza genetica altamente sensibili (2). È recentemente apparso un riesame della variazione biochimica all'interno e tra i membri del gruppo di specie Drosophila willistoni. È stata proposta una relazione diretta tra indici di somiglianza genetica e alcune designazioni sistematiche (3).
Se l'identità è considerata 1.0, è stato dimostrato che le popolazioni geografiche locali di una specie hanno somiglianze genetiche medie di 0.97, le sottospecie e le semispecie circa 0.8, le specie sorelle 0.52 e le specie non sorelle 0.35. Il presente documento riporta somiglianze genetiche all'interno e tra una coppia di specie parzialmente simpatiche, molto strettamente correlate, endemiche dell'Isola di Hawaii. Introduciamo l'uso di indici basati sulla variabilità cromosomica e li confrontiamo con indici basati sugli allozimi. La somiglianza biochimica sia all'interno che tra le specie è alta, sebbene siano fortemente differenziate morfologicamente e cromosomicamente. Queste specie sono evidentemente molto nuove nel senso biologico storico. Proponiamo che le loro somiglianze biochimiche siano direttamente correlate a questa circostanza.
Materiali e metodi
I campioni di Drosophila setosimentum e ochrobasis sono stati raccolti in foreste umide di alta montagna (1100-1600 m) sull'"Isola Grande" delle Hawaii, a cui sono endemici. Fatte salve le integrazioni di seguito, i dettagli delle località geografiche, della simpatria e delle tecniche citologiche ed elettroforetiche sono stati pubblicati (4). Nuove raccolte sono state effettuate a Puu Makaala, 8 km a nord-est di Olaa (località 11, Fig. 1 in rif. 4) e in due siti lungo il confine occidentale tra la Riserva Forestale di Kau e il Ranch Kahuku vicino all'estremità sud dell'Isola. Una di queste aree, chiamata Ranch Kahuku - 3800 ft. (1150 m), ha prodotto D. setosimentum. Altre nuove raccolte, per lo più di D. ochrobasis, sono state effettuate a Kipuka Pahipa, 7.2 km a nord-est del sito Ranch Kahuku - 3800 ft. (1150 m). Questo è lo stesso della località 14, descritta in rif. 4.
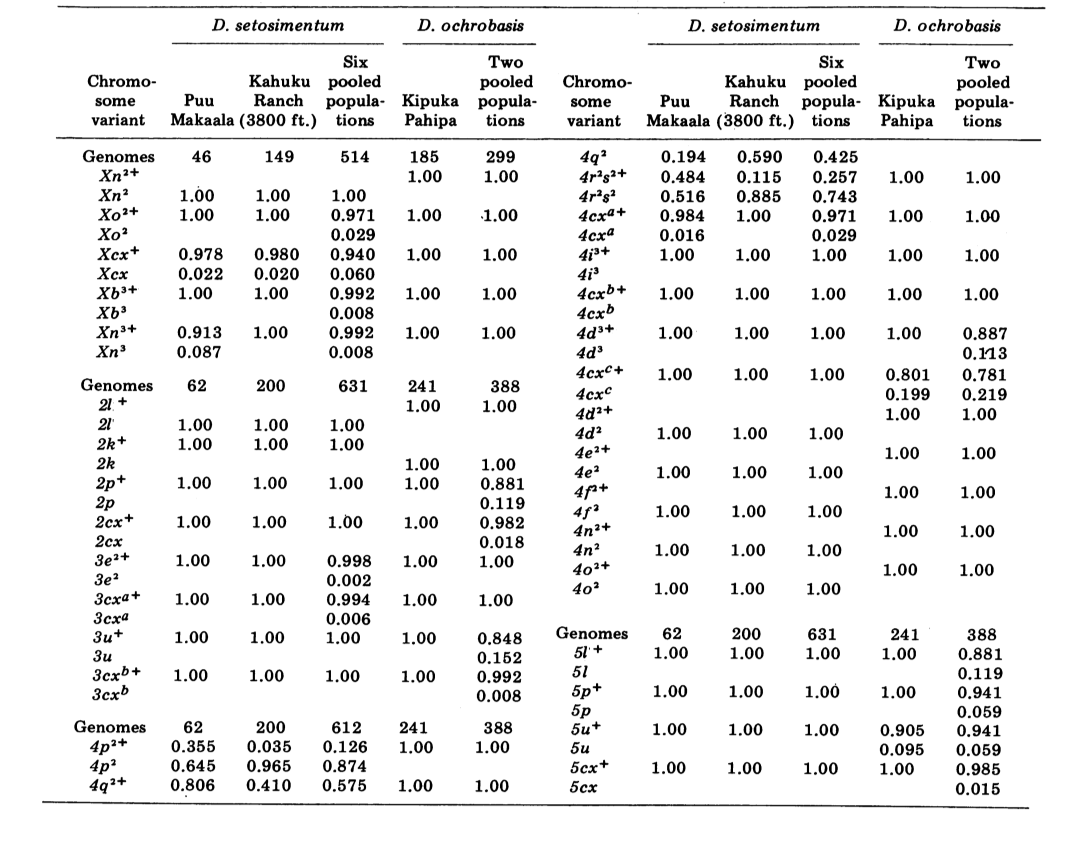
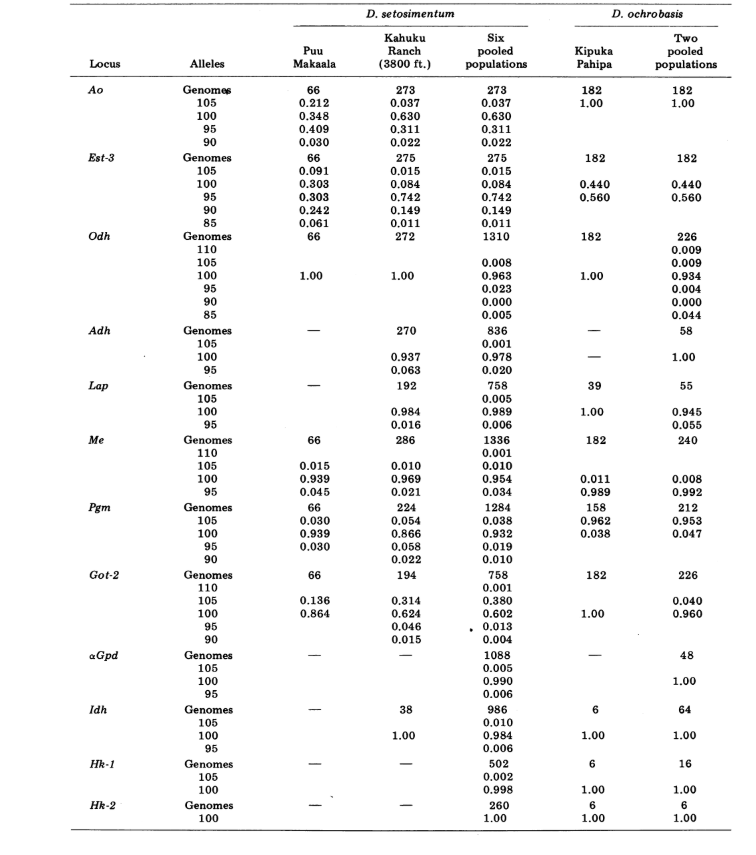
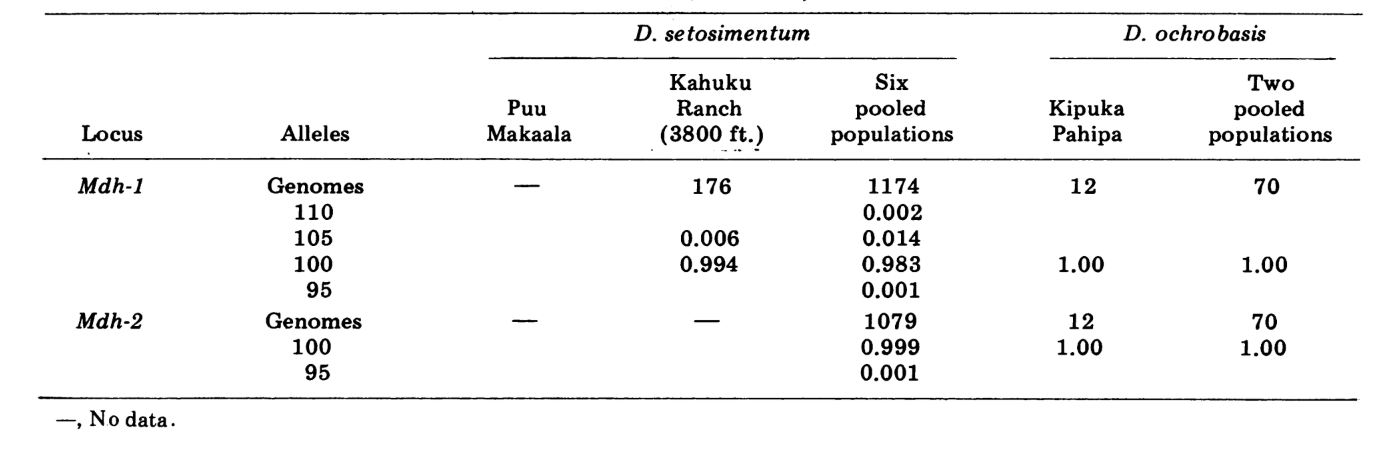
Riportiamo qui dati su tre ulteriori loci di allozimi, ossidasi dell'aldeide (Ao), un locus esterase β-naftil acetato (Est-3) e un ulteriore esochinasi (Hk-2), per un massimo di quattordici utilizzati per il confronto delle popolazioni. Le due specie mostrano variazione in un totale di 30 sezioni di cromosomi politenici a causa di inversioni o cromosomi complessi. Ogni sezione è stata valutata in ciascun striscio di cromosoma della ghiandola salivare esaminato; cioè, è stata registrata come eterozigote per la variante, omoziogote per la variante o omoziogote per la condizione standard. Questi dati sono stati utilizzati per calcolare i coefficienti di similarità nello stesso modo dei dati elettroforetici. La Tabella 2 elenca una nuova inversione, Xn3, da Puu Makaala. Il riesame di vecchie diapositive da Mawae, Kipuka-4400 (1340 m) ft. e Olaa mostra che la sua frequenza in queste popolazioni è rispettivamente 0.056, 0.032 e 0.005.
Osservazioni
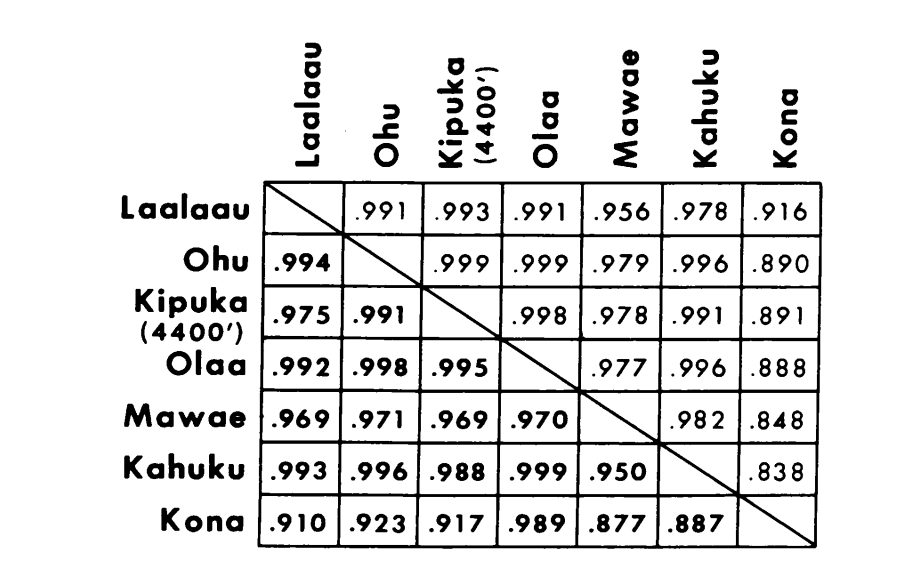
I nuovi dati e quelli di (4) sono stati utilizzati per calcolare i coefficienti di similarità (I di Nei) per entrambi gli allozimi (7-14 loci per confronto) e i cromosomi (30 sezioni per confronto), per sette popolazioni di D. setosimentum e tre di ochrobasis (Tabelle 1-3 e Fig. 1).
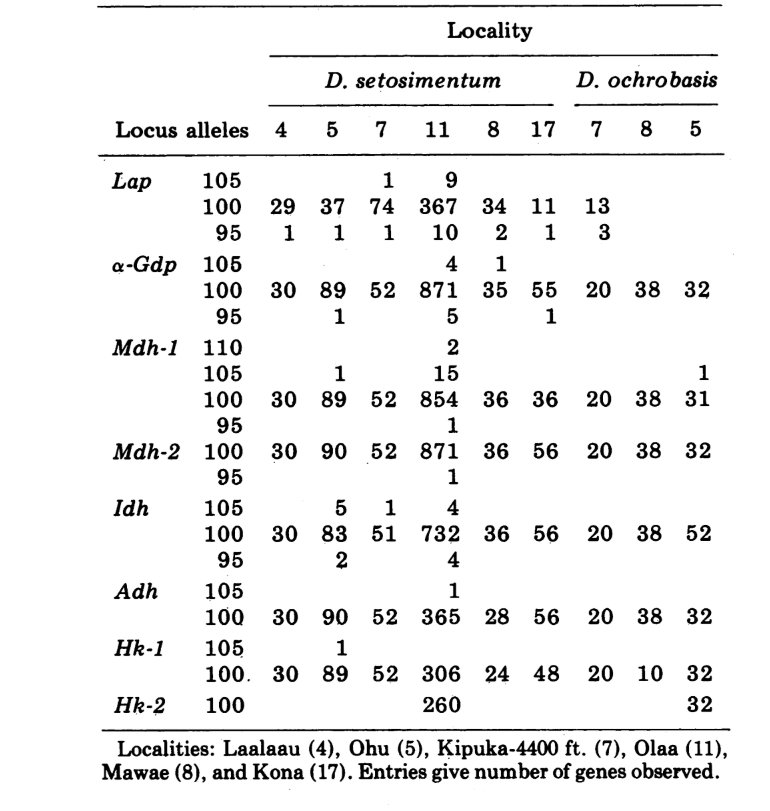
Le prime sei popolazioni di D. setosimentum elencate nella Fig. 1 provengono tutte dagli altopiani del lato sopravvento dell'isola. Mostrano coefficienti uniformemente elevati sia per i cromosomi che per gli allozimi e possono essere raggruppate. La media I con errore standard è indicata sulla riga superiore nel corpo della Tabella 4. La maggior parte dei confronti mostra alcune differenze rispetto alla popolazione di Kona, specialmente a livello cromosomico (Fig. 1 e Tabella 4). Quest'ultima è geograficamente isolata dalle altre sul lato sud-ovest dell'isola e presenta una differenza di eterocromatina metafasica fissa che non è inclusa nel calcolo dei coefficienti dei cromosomi politenici.
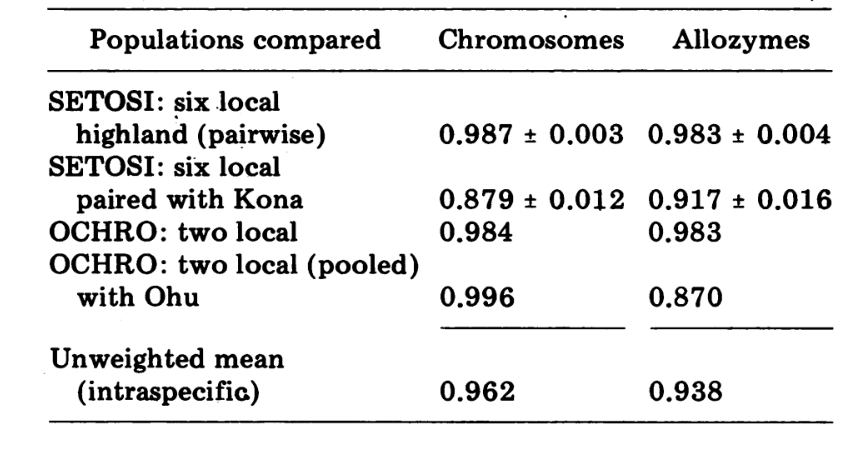
I dati di D. ochrobasis sono stati trattati in modo simile (Tabelle 1-5). L'esame a coppie delle due popolazioni, Kipuka Pahipa e i campioni combinati Mawae-Kipuka 4400 ft., mostra un'alta somiglianza (Tabella 4). Quando raggruppati, questi campioni mostrano una piccola differenza elettroforetica rispetto alla popolazione di ochrobasis di Ohu, che è isolata dalle altre sul vulcano Kohala nell'angolo nord-ovest dell'isola. Tuttavia, i valori medi intraspecifici non ponderati complessivi sono entrambi piuttosto alti e abbastanza simili (Tabella 4).
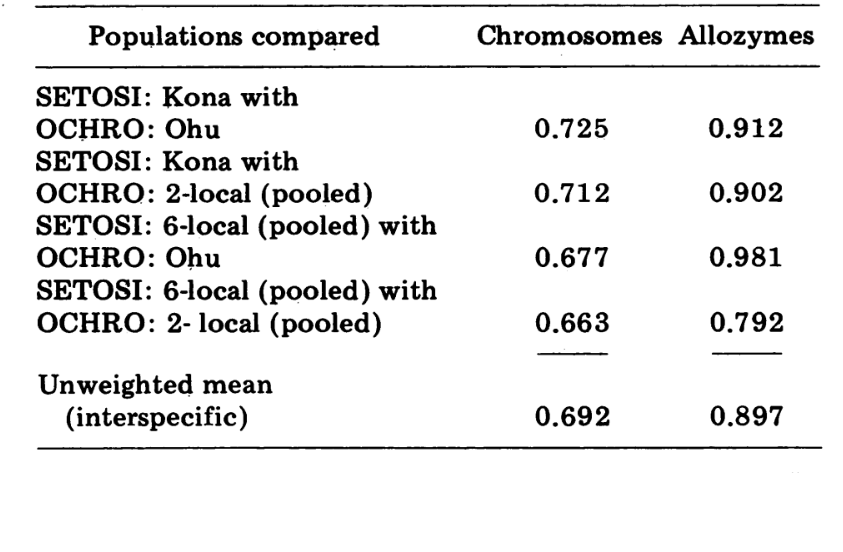
I confronti interspecifici sono mostrati nella Tabella 5. I confronti nella Tabella sono elencati in ordine di somiglianza cromosomica decrescente. I dati mostrano che i confronti cromosomici interspecifici sono molto più efficienti nel distinguere le specie rispetto a quelli basati sugli allozimi; infatti, tre dei quattro ultimi confronti sono superiori a 0.9. Il caso della popolazione Ohu di ochrobasis è interessante. Sebbene si distingua come diversa quando si confrontano gli allozimi all'interno della propria specie (0.870, Tabella 4), mostra una straordinaria somiglianza allozimica con setosimentum (0.981 con il pool di alta montagna e 0.912 con Kona, Tabella 5).
Discussione
Sebbene la differenziazione citologica tra queste due specie sia considerevole, le differenze biochimiche sono piccole. Tuttavia, le due entità non sembrano essere sottospecie. Gli individui di ciascuna possono essere chiaramente riconosciuti nella maggior parte delle situazioni simpatiche da una serie estesa di inversioni fisse e ci sono differenze altitudinali (4) e di siti di riproduzione (5) così come. I maschi delle due specie differiscono in modo sorprendente nel motivo di colore sulle ali (6).
D'altra parte, ci sono prove considerevoli che suggeriscono che D. setosimentum e ochrobasis potrebbero essere considerati semispecie (7). L'inferenza che ci sia stata una certa ibridazione tra di loro in passato (4) è stata ora confermata da una recente scoperta (8). Nella zona della Riserva Forestale Kahuku Raneh-Kau vicino all'estremità sud dell'isola, è stata inequivocabilmente riconosciuta una Fl naturalmente occorrente e tre ibridi backeross tra le specie. Questo corrisponde a circa il due per cento delle mosche selvatiche esaminate in quest'area. In considerazione dei campioni naturali sostanziali studiati e della ricchezza di marcatori sia elettroforetici (9) che citologici in quell'area, l'incrocio può essere giudicato piuttosto scarso.
Le differenze sessuali secondarie, come quelle che caratterizzano i maschi di queste specie, non si oppongono alla proposta di uno status di semispecie. Infatti, tali caratteri distinguono alcuni dei casi classici di semispecie, come, ad esempio, in certi uccelli del paradiso (7). Come negli uccelli, questi caratteri sessuali secondari riguardano il complesso comportamento territoriale (lek) e sessuale manifestato da molti del gruppo delle “Drosophila a ali a forma di immagine” a cui appartengono queste mosche (10).
Infine, il confronto elettroforetico più inclusivo (4° confronto, Tabella 5) dà un valore di 0.792, vicino a quello proposto per le semispecie di Drosophila paulistorum (3). A differenza di quest'ultima, tuttavia, D. setosimentum e ochrobasis sembrano essere meglio descritte come semispecie piuttosto avanzate, in cui l'isolamento riproduttivo in natura è ancora incompleto. Siamo favorevoli alla conservazione dei due nomi specifici per le due entità.
Rimane, tuttavia, il fatto interessante che alcune popolazioni delle due specie sono praticamente indistinguibili elettroforeticamente. La più estrema di queste è la popolazione Ohu di ochrobasis, che mostra un indice di somiglianza di 0.981 quando confrontata con D. setosimentum. Le informazioni citologiche parallele su questa popolazione escludono la possibilità che la somiglianza elettroforetica sia dovuta a ibridazione attuale.
Una stretta somiglianza elettroforetica interspecifica non è sconosciuta tra altre specie di Drosophila dell'Isola di Hawaii (11). Ad esempio, le specie simpatiche D. silvestris e D. heteroneura presentano ampie differenze morfologiche (12) e citologiche (13, 14). Mostrano isolamento etologico (15) così come altre differenze biologiche (16). Tutto ciò implica una considerevole differenza genetica oltre alle inversioni. Eppure, queste due specie mostrano un coefficiente di somiglianza allozimica di 0.96 (11). Secondo le proposte in (3), questo le collocherebbe come popolazioni locali della stessa specie, un'interpretazione insostenibile.
L'Isola di Hawaii è geologicamente nuova (meno di 7 x 105 anni). Le specie endemiche dell'Isola sono necessariamente ancora più nuove. Uno di noi (17) ha suggerito che la speciazione in queste e altre forme potrebbe aver coinvolto una riorganizzazione genetica forzata a seguito degli effetti stocastici di un evento fondatore allopatrico. A differenza degli oggetti di questo articolo, tuttavia, sono noti diversi casi in cui le specie si formano senza alcuna riorganizzazione dei cromosomi politenici rilevabile. È noto un gruppo di otto (ora 9) specie omosequenziali correlate a D. grimshawi (18). In modo simile, i casi attuali mostrano che le specie possono effettivamente essere formate anche con solo una piccola quantità di riorganizzazione allozimica. Suggeriamo che questo sia correlato alla recentezza degli eventi di speciazione. È possibile che le due popolazioni siano appena emerse da una fase fortemente stocastica nella loro storia vitale come specie. Questa fase potrebbe essere caratterizzata principalmente da una riorganizzazione ricombinazionale rispetto alla specie ancestrale e dall'istituzione di nuovi sistemi genetici internamente bilanciati. La fase di nuova risposta adattativa e tracciamento ambientale potrebbe essere appena iniziata.
Questa visione sottolinea anche un punto recentemente sollevato in una discussione sulle piuttosto sorprendenti somiglianze biochimiche tra l'uomo e lo scimpanzé (19). Gli eventi di speciazione possono coinvolgere geni regolatori che non sono normalmente rivelati dai metodi attuali nella genetica delle popolazioni.
Autori: H. L. Carson, W. E. Johnson, P. S. Nair, F. M. Sene